ת»ụ̀̉¼̉²ÏÉú²úº¬Ö©ÖëË¿µ°°×µÄĐÂĐ͸´ºÏ¼ëË¿ÏËάyd15135
ƠÅÔ¬ËÉ1,2£¬ƠỒ́¸£2£¬ƠÔ°®´º2£¬Îĺ́Đă2£¬ÖĐÔ«ÑÅĐÛ3 1.Î÷ÄÏ´óѧ·ÄÖ¯·₫װѧԺ£¬ÖØḈ400715£»2.¼̉²Ï»ụ̀̉×éÉúÎïѧ¹ú¼̉ÖصăʵÑéỂ(Î÷ÄÏ´óѧ)£¬ÖØḈ400715£»3.ÈƠ±¾ĐÅÖƯ´óѧÏËάѧ²¿£¬ÈƠ±¾³¤̉°ÏØ386-8567
ÊƠ¸åÈƠÆÚ£º2011-05-04 Đ̃»ØÈƠÆÚ£º2011-08-29
»ù½đÏîÄ¿£ºÖĐÑë¸ßĐ£»ù±¾¿ÆÑĐ̉µÎñ·ÑרÏî×ʽđ×ÊÖúÏîÄ¿(XDJK2009B007)£»½̀Óư²¿µÚ39ÅúÁôѧ»ù½đÏîÄ¿(½̀Íâ˾Áô[2010]1174ºÅ)£»½̀Óư²¿´ºêͼƻ®×ÊÖúÏîÄ¿(½̀Íâ˾Áô[2010]610ºÅ)£»ÈËÁ¦×ÊÔ´ºÍÉç»á±£Ơϲ¿ÁôѧÈËÔ±¿Æ¼¼»î¶¯ÓÅÏÈ×ÊÖú»ù½đÏîÄ¿(ÓåÈËÉç°́[2009]l16ºÅ)£»Î÷ÄÏ´óѧ²©Ê¿»ù½đÏîÄ¿(SWUB2007067)
×÷Ơß¼̣½é£ºƠÅÔ¬ËÉ(1971-)£¬ÄĐ£¬¸±½̀ÊÚ£¬²©Ê¿¡£ÑĐ¾¿·½Ị̈ΪĐÂĐÍÏËά²ÄÁÏÓ빦ÄÜ·Ä֯Ʒ¡£E-mail£ºyszhang@SWU.edu.en¡£
ÔÔØ£º·Ä֯ѧ±¨2012/5£»1-5
¡¾Ơª̉ª¡¿ÎªÁËÈ˹¤´óÁ¿Éú²úÖ©ÖëË¿µ°°×£¬Ö©ÖëÇ£̉ưË¿»ụ̀̉(MaSp1)±»ÓĂpiggyBacת»ụ̀̉ÔØ̀åµ¼Èë¼̉²ÏȾɫ̀åÖĐ¡£¼ëË¿µ°°×·ÖÎöÏÔʾת»ụ̀̉¼̉²Ï¿É̉ÔÉú²úº¬ÓĐÖ©ÖëÇ£̉ưË¿µ°°×µÄ¸´ºÏ¼ëË¿ĐÂÏËά¡£Óë̀́È»¼ëË¿Ïà±È£¬×ª»ụ̀̉¼̉²Ï¼ëË¿Ç¿¶ÈÔö¼Ó5.3£¥ £¬¶ø¶ÏÁÑÉ́³¤¼¸ºơĂ»ÓĐ±ä»¯¡£ÑĐ¾¿±íĂ÷ÀûÓĂת»ụ̀̉¼̉²Ï´óÁ¿Éú²ú¸÷ÖÖĐÂĐ͸´ºÏµ°°×ÖÊÏËά²ÄÁϾßÓĐDZÔÚµÄÓ¦ÓĂÇ°¾°¡£
¡¾¹Ø¼ü´Ê¡¿×ª»ụ̀̉¼̉²Ï£»Ö©ÖëË¿£»¸´ºÏ²ÏË¿
¡¾ÖĐͼ·ÖÀàºÅ¡¿TS 102.3£»Q 967 ÎÄÏ×±êÖ¾Â룺A ÎÄƠ±àºÅ£º0253-9721(2012)05-0001-05
Ö©ÖëÔÚÆäÉúĂüÆÚÖĐ¿É̉Ôͨ¹ư¸÷ÖÖÏÙ̀å²úÉúÈçÏÂ7ÖÖ²»Í¬µÄË¿µ°°×£º´óÄ̉×´ÏÙ²úµÄÇ£̉ưË¿(ÓÖĂûÇ£̉ưË¿¡¢ÍÏǣ˿)£»Đ¡Ä̉×´ÏÙ²úµÄ¿̣¼ÜË¿£»±̃×´ÏÙ²úµÄºáË¿£»¹Ü×´ÏÙ²úµÄÂѼëË¿£»¼¯ºÏÏÙ²úµÄºáË¿±íĂæđ¤ĐÔÎÀæ×´ÏÙ²úµÄ¸½×ÅÅ̀µ°°×£»ÆÏ̀Ñ×´ÏÙ²úµÄ²¶»ñË¿[1]¡£ƠâĐ©Ë¿µ°°×ÔÚÖ©Öë̀åÄÚË¿ÏÙÖĐÊÇ¿ÉÈÜĐԵĵ°°×ÖÊ£¬´óÄ̉×´ÏÙ·ÖĂÚµÄÇ£̉ưË¿̣̉Æ䳬¸ßµÄÇ¿¶ÈºÍµ¯ĐÔ¶ø±¸ÊܹØ×¢£¬Ç£̉ưË¿µÄ¶ÏÁÑÇ¿¶È³¬¹ưÁËͬµÈÖ±¾¶µÄ¸ÖË¿£¬¶ÏÁÑÉ́³¤Âʳ¬¹ư35£¥ £¬¸ßÓÚÆƠͨµÄ²ÏË¿ÏËά[2]¡£×÷Ϊ̀́È»ÉúÎï²ÄÁÏ£¬ÈËĂÇÏ£Íû»ñµĂÁ¦Ñ§ĐÔÄÜÓÅ̉́µÄÖ©ÖëË¿£¬½ø¶øÓ¦ÓĂÓÚ¹¤̉µ¡¢¾üÊ¡¢̉½̉©µÈÁ́Ọ́¡£Ö©ÖëË¿Ç£̉ưË¿ÏËάÓÉ2ÖÖÖ÷̉ªµ°°×Öʹ¹³É£¬·Ö±đÊÇMaSpl(major ampullate spidroin1)ºÍMaSp2µ°°×£¬ËüĂǹ¹³ÉÖ©ÖëË¿µÄ±ÈÀư´óÔ¼ÊÇ3:2¡£Ơâ2ÖÖµ°°×ÖÊ·Ö×ÓÁ¿Îª300 kDa×óÓ̉£¬ÆäºËĐIJ¿·ÖÊÇÓÉ100¸ö´®ÁªÖظ´Đ̣ÁĐ×é³É£¬Ă¿¸ö´®ÁªÖظ´Đ̣ÁĐÓÉ30¡«40¸ö°±»ùËá×é³É£¬ÆäÖи»º¬¸Ê°±Ëá¡¢±û°±ËáµÄ¸ß¶ÈÖظ´Đ̣ÁĐ£¬´óÁ¿µÄ¸Ê°±Ëá¡¢±û°±ËáÖظ´ÇøỌ́±»ÈÏΪĐγÉÖ©ÖëË¿ÏËάµÄ½á¾§Çø£¬ÎªÖ©ÖëÇ£̉ưË¿µÄ¸ßÇ¿Á¦×ö³ö¹±Ï×[3-4]¡£
ΪÁË»ñµĂ´óÁ¿µÄÖ©ÖëË¿µ°°×£¬ÑĐ¾¿ÈËÔ±½øĐĐÁ˸÷ÖÖ³¢ÊÔ£¬·Ö±đÔڴ󳦸˾ú¡¢½Íĸ¡¢Ö²Îï¡¢À¥³æϸ°û¡¢²¸È鶯Îïϸ°û¡¢É½Ṇ̃µÈË̃Ö÷½øĐĐÁËÖ©ÖëË¿µ°°×µÄ±í
´ï[5-8]£¬ÎÄÏ×[9]ÔÚ¼̉²Ïϸ°û¡¢Ó׳æÖĐ½øĐĐÁ˱í´ï£¬µ«ÊDzúÁ¿¶¼²»¸ß£¬½ö»ñµĂÉÙÁ¿¿ÉÈܽâĐÔµÄÖ©ÖëË¿µ°°×£¬̉ªĐγÉÏËά»¹±ØĐë¾¹ư»¯Ñ§·ÄË¿µÄ·½·¨£¬³É±¾·Ç³£¸ß¡£
¼̉²ÏÔÚ25 dµÄÓ׳æÉú³¤ÆÚÄÚ̀åÖØ¿ÉÔö¼Ó1Ị́±¶£¬ÆäÖĐÖ÷̉ªÊǽøĐе°°×Öʵĺϳɣ¬Ëù̉Ô¼̉²ÏÊǸßЧµÄµ°°×ÖÊÉúÎï·´Ó¦Æ÷£¬ÔÚË¿ÏÙÖĐºÏ³É´óÁ¿̉º̀¬µÄµ°°×ÖÊ£¬Í¨¹ưÍÂË¿Æ÷ÔÚ¿ƠÆøÖĐĐγɹ̀̀¬µÄ¼ëË¿¡£ÑĐ¾¿½á¹ûÏÔʾ£¬Ö©ÖëÇ£̉ưË¿(MaSp1)Óë¼̉²ÏµÄË¿ËØÖØÁ´(FibH)¾ßÓĐÏàËƵĸ߶ÈÖظ´ºËÜƠËáĐ̣ÁĐ£¬Æä°±»ùËá×é³É·Ç³£½Ó½ü[10]¡£²ÏºÍÖ©ÖëµÄÉúÎï·ÄË¿¹ư³̀̉²ÓĐĐ©ÏàËÆ£¬ËüĂǶ¼ÊÇÔÚ³£Î³£Ñ¹Ï½øĐеÄ̉º¾§·ÄË¿£¬¶øÇ̉¶¼ÄÜͨ¹ư·ÄË¿Æ÷¹Ù¿ØÖÆË¿µÄ½á¹¹ºÍ̀ØĐÔ¡£
½ü10ÄêÀ´£¬Đí¶àÑĐ¾¿̉Ѿ֤Ă÷piggyBacת»ụ̀̉ÔØ̀åÊÇ̉»ÖÖÏ൱ÓĐЧµÄ¼̉²Ïת»ụ̀̉¹¤¾ß£¬Æä·½·¨̉Ѿ±È½Ï³ÉẾ[11]¡£±¾ÎÄÀûÓĂpiggyBacÔØ̀彫֩ÖëË¿µÄÇ£̉ưË¿(MaSp1)»ụ̀̉µ¼Èë¼̉²ÏȾɫ̀壬̀½̀ÖÀûÓĂת»ụ̀̉¼̉²ÏÉú²úº¬ÓĐÖ©ÖëË¿µ°°×µÄĐÂĐ͸´ºÏ¼ëË¿ÏËά¡£
1 ʵÑ鲿·Ö
1.1 ²Ä ÁÏ
¼̉²ÏN4Æ·ÖÖ¡¢piggyBacת»ụ̀̉ÔØ̀åºÍMaSp1¿¹̀åÓÉÈƠ±¾ĐÅÖƯ´óѧÏËάѧ²¿̀ṩ£¬Ó׳æ25¡æ É£̉¶ËÇÓư¡£
1.2 ʵÑé·½·¨
1.2.1 ¿Ë¡¼̉²ÏË¿ËØÖØÁ´Æô¶¯×ÓºÍLBSĐ̣ÁĐ
¸ù¾Ư¼̉²ÏË¿Ëص°°×µÄÈ«»ụ̀̉Đ̣ÁĐÉè¼ÆÁË2¶ỔưÎï(primer)£¬·Ö±đÀ©Ôö¼̉²ÏË¿ËØÖØÁ´(FibH)Æô¶¯×ÓºÍLBS(L Chain Binding Site)Đ̣ÁĐ¡£
1.2.2 ת»ụ̀̉ÔØ̀å¹¹½¨
ÔÚ֮ǰµÄÑĐ¾¿ÖĐ£¬̉Ѿ»ñµĂÁË̉»¶Î1.5 kbµÄÑÇÖ̃Âçи¾Ö©Öë(Nephila clavata)Ç£̉ưË¿(MaSp1)DNAĐ̣ÁĐ[9]¡£Í¨¹ư»ụ̀̉¹¤³̀µÄ·½·¨£¬¹¹½¨ÁË̉»¸öÍêƠûµÄÔÚ¼̉²ÏË¿ËØÖØÁ´(FibH)Æô¶¯×Ó̉ưµ¼ÏµÄÖ©ÖëË¿Ç£̉ưË¿»ụ̀̉±í´ïĐ̣ÁĐ(ĂüĂûΪFibH-MaSp1-LBS)¡£
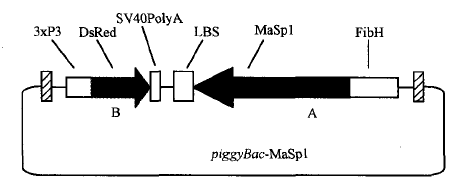
½«Ơâ¶ÎÍêƠû±í´ïĐ̣Áе¼ÈëpiggyBacÖÊÁ£¹¹½¨×ª»ụ̀̉ÔØ̀å(piggyBac MaSp1)£¬Èçͼ1ÖĐAËùʾ¡£
|
|
|
×¢£ºÁ½¶ËµÄ×óбÏß¿̣´ú±ípiggyBac-MaSplת×ù×ÓµÄ×óÓ̉²à̉íĐ̣ÁĐ£»3xP3ΪÈ˹¤ºÏ³ÉµÄ̉»¶ÎºËÜƠËáĐ̣ÁĐ£»SV40PolyAΪºï¿ƠÅƯ²¡¶¾40(SV40)µÄ¶à¾ÛÏÙÜƠËáĐ̣ÁĐ(PolyA)¡£ |
|
ͼ1 piggyBac-MaSplת»ụ̀̉ÔØ̀å½á¹¹ |
1.2.3 piggyBac-MaSplÖÊÁ£ÔØ̀åµÄÏÔ΢עÉä
½«piggyBac-MaSplÖÊÁ£ÓĂ³¬´¿Ë®µ÷Åä³É
0.4µg£¯mL£¬ÓĂÎÙƠëÏÈÔÚ²úÂÑ3¡«6 hµÄ¼̉²Ï²ÏÂÑ¿ª1¸öĐ¡¿×£¬È»ºóÓĂĂ«Ï¸²£Á§Ơ뽫ת»ụ̀̉ÔØ̀åpiggyBac-MaSplÖÊÁ£ÓëHelperÖÊÁ£̉»Æđ×¢Èë²ÏÂÑÖĐ£¬³£ÎÂÏÂÈĂ²ÏÂÑ·ơ»¯(GO´ú)¡£
1.2.4 ת»ụ̀̉¼̉²Ïɸѡ
ËÇÓưGO´ú¼̉²Ï£¬´ưÆ仯¶ê¡¢½»Åä¡¢²úÂѺ󣬶ÔG1´ú¼̉²Ï½øĐĐת»ụ̀̉ɸѡ¡£ÓĂÓ«¹âÏÔ΢¾µ¶Ô¼̉²ÏÓ׳桢Ӽ¡¢¶ê¸÷¸öÉú³¤Ê±ÆÚµÄÑÛ¾¦½øĐĐ¹Û²́£¬piggyBacת×ù×ÓĐ¯´øµÄº́É«±ê¼Çµ°°×(DsRed)»á·¢³öº́É«Ó«¹â£¬´Ó¶øɸѡ³öת»ụ̀̉³É¹¦µÄ¼̉²Ï¡£
1.2.5 ²Ï¼ëÖĐÖ©ÖëÇ£̉ưË¿µ°°×µÄ¼ø¶¨
½«ÆƠͨ²Ï¼ëºÍת»ụ̀̉¼̉²Ï²Ï¼ë¼ô³ÉËéƬ£¬ÓĂLiSCNÍêÈ«Èܽ⣬Ëù»ñµĂµÄµ°°×ÖÊÈÜ̉ºÓĂSDS.polyacrylamide gel electrophoresis(PAGE)½øĐеçÓ¾¼́²â£¬È»ºóÓĂĂẩß·´Ó¦(West blotting)ÑéÖ¤²Ï¼ëÖĐ´æÔÚÖ©ÖëË¿µ°°×¡£
1.2.6 ¸´ºÏ¼ëË¿ÏËάµÄÁ¦Ñ§ĐÔÄܲⶨ
Ëæ»úÑ¡È¡20Á£ÆƠͨ²Ï¼ëºÍת»ụ̀̉¼̉²Ï²Ï¼ëΪ²ÄÁÏ£¬¶Ôµ¥¸ù¼ëË¿ÏËά½øĐĐÇ¿Á¦ºÍ¶ÏÁÑÉ́³¤ÂʵIJⶨ¡£²âÊỔÇÆ÷²ÉÓĂUTM-I11-100ĐÍÇ¿Á¦̉Ç(ÈƠ±¾¶«ÑóBaldwinÓĐÏ̃¹«Ë¾)¡£
2 ½á¹ûÓë̀ÖÂÛ
2.1 ת»ụ̀̉ÔØ̀åµÄÉè¼Æ
±¾ÎÄÉè¼ÆµÄת»ụ̀̉ÔØ̀åÈçͼ1Ëùʾ¡£Ë¿ËØÖØÁ´Æô¶¯×Ó(Fibroin
H promoter)̉ưµ¼Ơû¸öÖ©ÖëË¿»ụ̀̉ÔÚË¿ÏÙϸ°û±í´ï£¬ÖØÁ´LBS(L Chain Binding Site)Đ̣ÁĐÈĂÖ©ÖëË¿MaSp1µ°°×ÓëÇáÁ´µ°°×½áºÏ£¬Í¬Ê±LBSĐ̣ÁĐÄ©¶ËÓĐpolyAĐ̣ÁĐ£¬Ê¹Ơû¸öÖ©ÖëË¿µÄ±í´ïÖƠÖ¹£¬ƠâÑù¾ÍÈ˹¤ºÏ³ÉÁË̉»¸öÍêƠûµÄ±í´ïµ¥Ôª£¬Ơâ¸ö±í´ïµ¥ÔªÇ¶Èëµ½piggyBacת»ụ̀̉ÔØ̀åµÄת×ù×Ó²¿·Ö(¼ûͼlÖĐA)¡£
ÔÚת×ù×Ó²¿·Ö»¹ÓĐ̉»¸öÍêƠûµÄº́É«Ó«¹âµ°¾Ê(DsRed)µÄ±í´ïµ¥Ôª(¼ûͼl ÖĐ B)£¬ËüÓÉ3xP3Æô¶¯×Ó̉ưµ¼£¬SV40PolyAÖƠÖ¹±í´ï¡£3xP3Æô¶¯ÓèÖ»ÔÚÑÛϸ°ûÖĐÆô¶¯ºóĐø»ụ̀̉µÄ±í´ï£¬Ëù̉ÔDsRed×÷Ϊ±ê¼Ç»ụ̀̉ÔÚ¼̉²Ï¸÷¸öÉú³¤Ê±ÆÚÖ»ÔÚÑÛϸ°ûÖбí´ïDsRedÓ«¹âµ°°×.ÔÚÓ«¹âÏÔ΢¾µÏ¿É̉Ô¿´µ½¼̉²ÏÑÛ¾¦·¢³öº́É«Ó«¹â£¬ƠâÊÇ̉»¸ö±ê¼Çµ°°×(Marker)£¬¿É̉Ô×÷Ϊת»ụ̀̉³É¹¦µÄ̉À¾Ư[11]¡£
ÑĐ¾¿±íĂ÷£¬¼̉²ÏµÄË¿ËØÖØÁ´(H Chain)µ°°×C-Ä©¶ËµÄ°ëë×°±Ëá±ØĐëÓëÇáÁ´(L Chain)µÄ°ëë×°±Ëáͨ¹ư¶₫Ạ́¼ü½áºÏÔÚ̉»Æđ.²ÅÄÜĐγÉÍêƠûµÄË¿Ëص°°×[12]¡£ÓÉÓÚ±¾ÎIJÉÓĂ¼̉²ÏµÄË¿ËØÖØÁ´µ°°×(FibH)Æô¶¯×Ó̉ưµ¼Ö©ÖëË¿µ°°×µÄ±í´ï£¬Ëù̉Ô±ØĐëÔÚÖ©ÖëË¿µ°°×»ụ̀̉Đ̣ÁĐºóĂæÁ¬½ÓË¿Ë÷ÖØÁ´»ụ̀̉Ä©¶ËµÄLBSµ°°×»ụ̀̉Đ̣ÁĐºóĂæÁ¬½ÓË¿ËØÖØÁ´»ụ̀̉Ä©¶ËµÄLBSĐ̣ÁĐ£¬¸ĂĐ̣ÁĐº¬ÓĐË¿Ë÷ÖØÁ´C-Ä©¶ËµÄ°ëë×µªËáºÍʹ˿ËØÖØÁ´µ°°×±í´ïÖƠÖ¹µÄpolyAĐ̣ÁĐ£¬ƠâÑù²ÅÄܹ¹½¨̉»¸öÓÉÖ©ÖëË¿»ụ̀̉´ú̀æË¿ËØÖØÁ´»ụ̀̉µÄÍêƠû±í´ïĐ̣ÁĐ£¬Ëù±í´ïµÄÖ©ÖëË¿µ°°×²ÅÄܹ»ÔÚ¼̉²ÏË¿ÏÙϸ°ûÖĐÓëÇáÁ´µ°°×½áºÏ¡£·ÖĂÚµ½Ë¿ÏÙ¹ÜÇ»ÖĐĐγɼëË¿µÄË¿Ëز¿·Ö¡£
2.2 ת»ụ̀̉ЧÂÊ
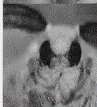
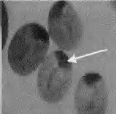
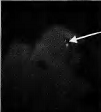
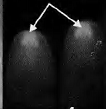
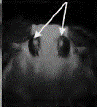
ÏÔ΢עÉäÁË1908Á£²ÏÂÑ£¬ÓĐ148Á£·ơ»¯³öGO´úÓ׳æ,GO´úÓ׳澹ư»¯Ó¼¡¢»¯¶ê¡¢½»Åäºó²ú³öG1´ú²ÏÂÑ56¸ö¶êȦ£¬Gl´ú²ÏÂÑס·ơ»¯Ç°3-4 d¿ªÊ¼ÓĂÓ«¹âÏÔ΢¾µ¹Û²́δ·ơ»¯¼̉²ÏÓ׳æµÄÑÛ¾¦(¼ûͼ2ÖĐA)£¬·¢³öº́É«Ó«¹âµÄÑôĐÔÓĐ2¸ö¶êȦ,̀ô³öÑôĐÔ¸ö̀å¼̀ĐøËÇÓư¡£
ת»ụ̀̉ЧÂÊÈç±ílËùʾ£º
±íl ת»ụ̀̉ЧÂÊ
|
ÏîÄ¿ |
Á£Êư |
|
G0´ú×¢Éä²ÏÂÑÊư |
1908 |
|
G0´ú·ơ»¯²ÏÂÑÊư |
148 |
|
Gl´ú²úÂѶêȦÊư |
56 |
|
G1´úÑôĐÔ¶êȦÊư |
2 |
|
G1´úÑôĐÔÂÊ£¯£¥ |
3.57 |
ÆäG0´ú·ơ»¯ÂÊÖ»ÓĐ7.8£¥¡£Óëδ½øĐĐÏÔ΢עÉäµÄÆƠͨ¼̉²Ï·ơ»¯Ïà±È.ÏÔ΢עÉä¶Ô²ÏÂÑỐ³É̉»¶¨³̀¶ÈµÄËđÉË¡£·ơ»¯ÂÊĂ÷ÏÔ½µµÍ¡£×ʼÓÉÓÚ¿ª¿×¹ư´ó,ÉơÖÁ³öÏÖG0´úÍêÈ«²»·ơ»¯µÄÇé¿ö£¬Ëù̉Ôת»ụ̀̉ЧÂÊÓë×¢Éä²Ù×÷µÄẾÁ·³̀¶ÈĂÜÇĐÏà¹Ø¡£GI´úÑôĐÔ¸ö̀åÓĐ8Í·Ó׳æ(·ÖÊô2¸ö¶ÈĂÜÇĐÏà¹Ø£¬Gl´úÑôĐÔ¸ö̀åÓĐ8Í·Ó׳æ(·ÖÊô2¸ö¶êȦ),ÑôĐÔÂÊΪ3.57%¡£ÔÚ²ÏÂÑ¡¢Ó׳桢Ӽ¡¢¶êµÈ²»Í¬Ê±ÆÚ£¬¶¼¹Û²́µ½ÑÛ¾¦·¢³öÎȶ¨µÄº́É«Ó«¹â(¼ûͼ2(b))¡£
|
|
A |
B |
C |
D |
|
a.¹âѧÏÔ΢¾µƠƠƬ |
|
|
|
|
|
b.Ó«¹âÏÔ΢¾µƠƠƬ |
|
|
|
|
|
ͼע£ºA-²ÏÂÑ£¬B-5ÁäÓ׳棬C-²ÏÓ¼£¬D-²Ï¶ê ͼ£¨b£©ÖĐ°×É«¼ưÍ·Åúʾ·¢Ó«¹âµÄÑÛ¾¦²¿Î» |
||||
|
ͼ2 ת»ụ̀̉¼̉²Ï²»Í¬Ê±ÆÚµÄÓ«¹â¼́²â |
||||
2.3 ²Ï¼ëµ°°×ÖÊ·ÖÎö
ÆƠͨ¼̉²ÏºÍת»ụ̀̉¼̉²ÏµÄ²Ï¼ëÓĂLiSCNÈܽâºó½øĐĐSDS¡¤PAGEµ°°×ÖÊ·ÖÎö.½á¹ûÈçͼ3(a)Ëùʾ¡£
|
|
|
(a)SDS-PAGE
(b)Ăẩß·´Ó¦ |
|
ͼ3 ²Ï¼ëµ°°×ÖÊ·ÖÎö |
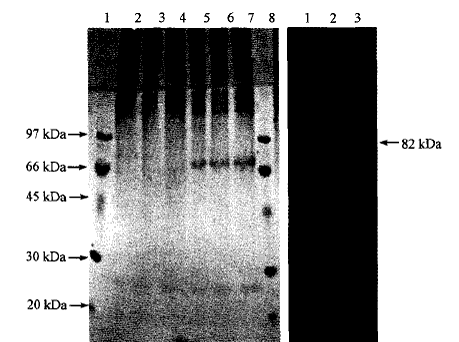
ͼ3(a)ÖĐl¡¢8ΪµÍ·Ö×ÓÁ¿±ê¼Çµ°°×(LWM)£¬2¡«4ΪÆƠͨµÄN4¼̉²Ï²Ï¼ëÑùÆ·£¬5¡«7Ϊת»ụ̀̉¼̉²Ï²Ï¼ëÑùÆ·.ÔÚת»ụ̀̉¼̉²Ï²Ï¼ëÖĐÓĐ̉»¸ö82 kDaµÄµ°°×̀ơ´ø£¬ƠâÓë¼ÆËăµÄÖ©ÖëË¿MaSplµ°°×ÖÊ·Ö×ÓÖÊÁ¿̉»Ö£¬ÆƠͨµÄ¼̉²Ï²Ï¼ëÖĐĂ»ÓĐƠâ¸öµ°°×̀ơ´ø¡£
ÓẰ́È»MaSp1µ°°×¿¹̀å½øĐĐĂẩß·´Ó¦.½á¹ûÈçͼ3(b)Ëùʾ¡£×ª»ụ̀̉¼̉²ÏµÄºó²¿Ë¿ÏÙ(posl silkgland£¬PSG)ºÍ²Ï¼ëµ°°×ÑùÆ·¶¼ÔÚÏàÓ¦µÄ82 kDaλÖĂ³öÏÖÁËÖ©ÖëË¿MaSplµ°°×Ăẩß·´Ó¦̀ơ´ø(1¡¢3)£¬¶øÆƠͨ²Ï¼ëÔÚ¸ĂλÖĂĂ»ÓĐ·´Ó¦(2)¡£Ơâ˵Ă÷ÔÚת»ụ̀̉¼̉²Ï²Ï¼ëÖĐ³É¹¦µØ±í´ïÁËÖ©ÖëË¿Ç£̉ưË¿µ°°×(MaSp1)¡£×ª»ụ̀̉¼̉²Ï·ÖĂڵļëË¿ÊÇ̉»ÖÖº¬ÓĐÖ©ÖëÇ£̉ưË¿µ°°×µÄĐÂĐ͸´ºÏ¼ëË¿¡£
2.4 ¸´ºÏ¼ëË¿Á¦Ñ§ĐÔÄÜ
¶ÔÆƠͨ²Ï¼ëºÍת»ụ̀̉²Ï¼ëµÄ¼ëË¿½øĐĐÁ¦Ñ§ĐÔÄܶԱȲâÊÔ£¬¼ÆËăƽ¾ùÖµ£¬½á¹ûÈçͼ4Ëùʾ¡£
|
|
|
ͼ4 ¸´ºÏ¼ëË¿ÏËάÁ¦Ñ§ĐÔÄÜ |
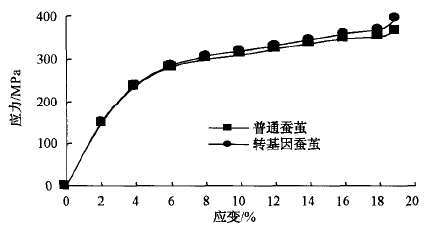
ÆƠͨN4Æ·ÖÖÚ£²Ï¼ë˿ƽ¾ùÇ¿¶ÈΪ376 MPa£¬¶ÏÁÑÉ́³¤ÂÊΪ19.2£¥ £»×ª»ụ̀̉¼̉²Ï¼ë˿ƽ¾ùÇ¿¶ÈΪ396 MPa£¬¶ÏÁÑÉ́³¤ÂÊΪ19£¥ ¡£×ª»ụ̀̉¼̉²Ï¼ëË¿Ç¿¶ÈÔö¼Ó5.3£¥ £¬¶ø¶ÏÁÑÉ́³¤Âʼ¸ºơĂ»ÓĐÇø±đ¡£
¸ßĐÔÄܵÄÖ©ÖëË¿ÏËά×÷ΪÉúÎï²ÄÁÏ¿É̉ÔÓ¦ÓĂÓÚ¹¤̉µ¡¢º½¿Ơ¡¢̉½ÁƵÈÁ́Ọ́£¬È»¶øÓÉÓÚÖ©Ö뼯ÖĐËÇÑøÓĐ×ÔÏà²ĐɱµÄÏ°ĐÔ£¬Ëù̉ÔÎ̃·¨Ïñ¼̉²Ï̉»Ñù´ó¹æÄ£È˹¤ËÇÑø£¬ÔÚ×ÔÈ»½çÖĐ²É¼¯»áʹ³É±¾¹ư¸ß¡£̣̉´Ë£¬ÔËÓĂÉúÎ﹤³̀µÄ·½·¨´óÁ¿Éú²úÖ©ÖëË¿µ°°×¾Í³ÉÁËÍ»ÆÆƠẩ»Æ¿¾±µÄ×î¼ÑÑ¡Ôñ¡£¶øÀûÓĂ¼̉²ÏÉú²úº¬Ö©ÖëË¿µ°°×µÄ¸´ºÏ¼ëË¿ÏËά²ÄÁϾßÓжÀ̀صÄÓÅÊÆ£º¿É̉Ô¹æÄ£»¯ËÇÑø£¬³É±¾µÍÁ®µÈ[9]¡£±¾ÎĽ«MaSp1»ụ̀̉ÓÉFibHÆô¶¯×Ó̉ưµ¼£¬Í¨¹ưpiggyBacת×ù×ÓƠûºÏ½ø¼̉²ÏȾɫ̀åÖĐ£¬ÊµÑéÖ¤Ă÷Ö©ÖëË¿µ°°×ÔÚת»ụ̀̉¼̉²ÏµÄ¼ëË¿Öеõ½±í´ï¡£
¼̉²Ï¼ëË¿ÓÉ75£¥Ë¿ËغÍ20£¥ Ë¿½º×é³É£¬ÆäÖĐË¿ËØÓÉÖØÁ´µ°°×(FibH)ºÍÇáÁ´µ°°×(FibL)×é³É£¬Đí¶àÑĐ¾¿ÓĂFibH¡¢FibLºÍË¿½º(sericin)µ°°×ÖʵÄÆô¶¯×ÓÔÚ¼̉²ÏË¿ÏÙÖбí´ïÍâÔ´µ°°×[13-16]¡£±¾ÎÄÑĐ¾¿ÏÔʾFibHÆô¶¯×Ó̉»Ñù¿É̉ÔÓĐЧ̉ưµ¼Ö©ÖëË¿MaSplµ°°×µÄ±í´ï¡£Í¬Ê±£¬ÓĂ΢ÉúÎɽṆ̃¡¢Ñ̀²ƯµÈÆäËûË̃Ö÷Éú²úµÄÖ©ÖëË¿µ°°×Đè̉ª̉»¸öÈ˹¤·ÄË¿µÄ¹ư³̀Éú³ÉÏËά£¬¶øÀûÓĂ¼̉²ÏË¿ÏÙÉú²úÏËά²ÄÁϾßÓжÀ̀صÄÓÅÊÆ£¬Ëü¿É̉Ôͨ¹ư¼̉²ÏµÄÉúÎï·ÄË¿¹ư³̀Ö±½ÓĐγÉÏËά¡£
Ä¿Ç°¸´ºÏ¼ëË¿µÄÁ¦Ñ§ĐÔÄÜĂ»ÓĐ´óµÄ¸ÄÉÆ£¬Ơâ¿ÉÄÜÓ븴ºÏ¼ëË¿ÖеÄÖ©ÖëË¿µ°°×±ÈÀư¹ưµÍÓĐ¹Ø(´óԼΪ3£¥)£»Í¬Ê±Ö©ÖëË¿Ö÷̉ªÓÉMaSp1ºÍMaSp2Ơâ2ÖÖµ°°×ÖÊ×é³É£¬ËüĂÇ̉ÔÊ²Ă´ÑùµÄ·½Ê½½áºÏ¹²Í¬ÎªÖ©ÖëË¿µÄ¸ßĐÔÄÜ×ö¹±Ï×£¬»¹ÓĐ´ưÑĐ¾¿¡£½ñºó¿É̉Ô´ÓÑĐ¾¿Ö©ÖëË¿µ°°×Óë²ÏË¿µ°°×µÄ±ÈÀư¡¢Ï໥¹ØϵÈëÊÖÉîÈëÑĐ¾¿Ơâ2ÖÖµ°°×ÖÊÔÙÉúĐγɵÄĐÂÏËά²ÄÁϵÄĐÔÄܺͻúÖÆ¡£
3 ½áÂÛ
ÀûÓĂpiggyBacת»ụ̀̉ÔØ̀壬½«¼̉²ÏÖØÁ´Æô¶¯×Ó(FibH promoter)̉ưµ¼ÏµÄÖ©ÖëÇ£̉ưË¿MaSp1µ°°×±í´ïµ¥Ôªµ¼Èë¼̉²ÏȾɫ̀åÖĐ£¬µĂµ½×ª»ụ̀̉³É¹¦µÄ¼̉²Ï¡£Í¨¹ưĂẩßʵÑéÖ¤Ă÷ת»ụ̀̉¼̉²Ï·ÖĂÚÁ˺¬ÓĐÖ©ÖëË¿µ°°×µÄ¸´ºÏ¼ëË¿¡£¾¡¹Ü¸´ºÏ¼ëË¿µÄÁ¦Ñ§ĐÔÄÜĂ»ÓĐĂ÷ÏÔ¸ÄÉÆ£¬µ«ÑĐ¾¿±íĂ÷£¬ÀûÓĂת»ụ̀̉¼̉²Ï´óÁ¿Éú²ú¸÷ÖÖĐÂĐ͸´ºÏµ°°×ÖÊÏËά²ÄÁϾßÓĐDZÔÚµÄÓ¦ÓĂÇ°¾°¡£
²Î¿¼ÎÄÏ×£º
[1] RISING A£¬NIMMERVOLL H£¬GRIP S£¬et a1.Spider silk protein mechanical property and gene sequence[J]Zool Sci£¬2005£¬22£º273-283
[2] Hu x£¬VASANTHAVADA K.KOHLER K.et.a1.
Molecular mechanisms of spider silk[J]Cell Mol Life Sci.2006£¬63£º1986-1999
[3] XU M,LEWIS R V. Structure of protein superfiber spider dragline silk[J].Proc Natl Acad Sci USA£¬1990£¬87£º7120-7124.
[4]HIMAN M B. LEWIS R V. Isolation of a clone
encoding a second dragline silk fibroin Nephila clavipes dragline silk is a two-protein fiber[J].J Biol Chem£¬1992£¬267£º19320-19324.
[5]FAHNESTOCK S R£¬IRWIN S L. Synthetic spider dragline silk proteins and their production in Escherichia coli[J].Appl Microbiol Biotechnol£¬1997£¬47£º23-32.
[6]FAHNESTOCK S R£¬ Bedzyk L A. Production of synthetic spider dragline silk protein in Pichia pastoris[J]. Appl Microbiol Biotechnol£¬1 997£¬47£º33-39.
[7]SCHELLER J£¬ GUHRS K H £¬ GROSSE F£¬ et a1.Production of spider silk proteins in tobacco and potato[J].Nat Biotechnol£¬2001£¬19£º573-577.
[8]TEULE F£¬ COOPER A R£¬ FURIN W A £¬ et a1. A protocol for the production of recombinant spider silklike proteins for artificial fiber spinning[J].Nat Protoc£¬2009(4)£º341-355.
[9]ZHANG Y£¬ HU J£¬ MIAO Y£¬ et a1. Expression of EGFP-spider dragline silk fusion protein in BmN cells and larvae of silkworm showed the solubility is primary limit for dragline proteins yield[J]. Mol Biol Rep£¬2008£¬35£º329-335.
[10]ZHANG Y£¬ SHIMIZU K£¬ SHIOMI K£¬ et a1. cDNA cloning of Nephila clavata dragline silk(MaSp1)gene and comparison with the sequence of Bombyx mori fibroin heavy chain[J].Sanshi.Konchu Biotec£¬2008£¬77£º39-45.
[11]TAMURA T£¬THIBERT C£¬ROYER C£¬et a1. Germline transformation of the silkworm Bombyx mori L. using a piggyBae transposon¡ªderived vector[J].Nat Biotechno1.2000£¬18£º81-84.
[12] TANAKA K£¬INOUE S£¬MIZUNO. SHydrophobic interaction of P25£¬containing Asn-linked oligosaccharide chains. with the H-L complex of silk fibroin produced by Bombyx mori[J].Insect Biochem Mol Biol£¬1999£¬29£º269-276.
[13]¶Î½¨Æ½£¬Đ́ºº¸££¬ÂíÈưÔ«£¬µÈ.ÈËÄÔÔ´ĐÔÉñ¾ÓªÑø̣̉×Ó»ụ̀̉(hBDNF)ÔÚת»ụ̀̉¼̉²ÏË¿ÏÙÖеÄ̀Ø̉́±í´ï[J].²Ï̉µ¿Æѧ£¬2009£¬35(2)£º248-252.
[14]Àî´º·å£¬»Æ¿Æ£¬¸Ê½ø·æ£¬µÈ.תֲËáø»ụ̀̉¼̉²ÏµÄÖÆ×÷¼°±í´ï¼́²â[J].À¥³æѧ±¨£¬2009£¬52(7)£º713-720.
[15]TOMITA M £¬HINO R£¬OGAWA S£¬et a1. A germline transgenic silkworm that secretes recombinant proteins in the sericin layer of cocoon[J].Transgenic Res£¬2007£¬I6£º449 -465.
[16]ZHAO A£¬ZHAO T£¬ZHANG Y£¬et a1.New and highly eficient expression systems for expressing selectively foreign protein in the silk glands of transgenic silkworm[J].Transgenic Res£¬2010£¬19(1)£º29-44