����ά���������֯Ʒ��ø����yd14114
������ �Ϻ���֯��ѧ�о�Ժ�����ݣ�
��� ȫ��Ⱦ���¼���Ӧ���ƹ�Э����
ԭ�أ�ȫ��Ⱦ���¼���Э������Ѷ2011��8/10/12
����֯ƷȾ���ӹ��У��˽���������Ư��ǰ����������Ӱ��ӡȾ��Ʒ�����Ĺؼ����ݳƣ�ӡȾ��Ʒ�ò�����80%���ɲ�����ǰ������ɵġ��ڴ�ͳ��ǰ���������У�ǿ�ǿ�ᡢ�������Ȼ�ѧҩ�����ڸ��ºͳ�ʱ��ľ��Ҽӹ������£�������ά���ܻ���ɺܴ����ˣ������ô�����ˮ����Դ�����ŷŵķ�ˮȾ�۳̶����أ�ƽ��CODԼΪ3000~3500mg/L��������ԼռȾ���ӹ����ŷ�����60%�������ϣ�����Щ�ѳ�ΪĿǰȾ����ҵ������չ����Ҫ�ϰ���������Ե�����ø�Ƽ����з�֯Ʒ��ǰ��������Ŀǰ�������ע�ĸ�Ч���ͺġ������ɫ�ӹ������ǽ��Ⱦ����ҵ��һ����Ҫ��չ�����������ǽ��Ķ��й����ϵ�ժ¼�������ĵģ����Ķ������У�������ﻯѧ���������ѧ����ѧ�Ƶ���º����Ϻ����̺���Ҳ�����Լ�֪ʶ��ƶ�������Ŀ��ܹ�һ©��ּ������ɳ���������ڴ����ߵ����������ָ�������ڴ�ø�Ƽ���ѧ�Һ�Ⱦ���������ܶԴ��и���Ĺ�ע�ͽ��ܺ�����������һ��ո�µĸ߿Ƽ�����ɫȾ����ҵ���츣�����ࡣ
һ������ά�Ľṹ�����
��һ������ά����������
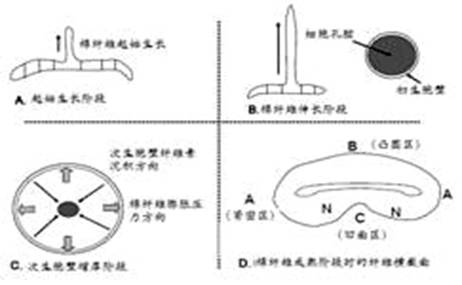
����ά�������ĸ��Σ���ͼ1��ʾ��
��1����ʼ�����Σ�ͼ1�е�A�������꿪����2�죬����ά��ʼ���������ѵ����Ƥͻ�� �γɹ���״����ά��ϸ����
��2������ά�쳤�Σ�ͼ1�е�B������20-25���У�ͻ��Ĺ���״����άϸ�����㣨�����ͳ������ڣ����ϵ������쳤���γ��������ά�ij��ȡ�
��3��������������Σ�ͼ1�е�C�����ڿ������35��50��ʱ���ڣ���ά�ش���Ӳ��ϵ���ϸ�����������ij������γɴ������ڣ���ʹ����άϸ��������������ʱ����������άϸ�����㣨�����ͳ������ڣ����൱ǿ�Ⱥ���(��)�ԣ��������������ѹ����Ϊ�������������ṩ�ռ䡣
��4������Σ�ͼ1�е�D����ǰ�������ζ����ڷ�յĹ����н��еġ�����ɴ�����������κ��εĹ��������ѣ��ڲ�ˮ�����������ڴ�����������ά���ܶȲ�ͬ�������������У�����ά���ν�������ѹ�����������Σ��γ�ͼ�е�A(������)��B��Բ������C������������N�����������ĸ��������ǵĿɼ��ԣ�Accessibility��˳����N > C > B > A��
|
|
|
ͼ1 ����ά��������ʾ�� |
����������ά�Ľṹ�Ͱ��������
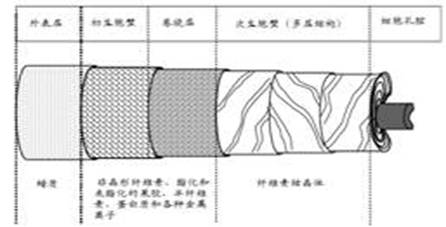
����ά�Ƕ��ͬ���ĵ�ϸ����֯��������㡢�������ڡ����Ʋ㡢�������ں�ϸ����ǻ��ɡ�����ͼ��
|
|
|
ͼ2 ����ά�Ľṹ |
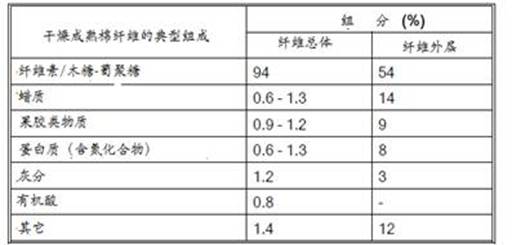
�ڳ�������ά�Ļ�ѧ����У�����90%��������ά���⣬�����൱���Ĺ����ʡ�����������ά�غͺ������ʵȰ�������1��ʾ
��1 ����ά�Ļ�ѧ���
|
|
|
|
����������ά������㣨cuticle��
����ά�����������һЩ��ˮ�Ե���ľ֬�ͽ����ص������������ɵı���⻬�ı�Ĥ������ֻ��12 nm�����ṹ���ܣ���ֲ�����������У�������Ȼ�������¶ȡ�ˮ����ʧ�ͳ溦�ȣ������˺������������㡣����ϰ�߰�����ά����Щ֬�����ʳ�Ϊ������cotton waxes�������������۵�ܸߣ���ΧҲ�ܹ㣨64��C - 214��C������ɺܸ��ӣ��京��������ά���ص�0.4 -0.8%������ˮ������С��������ά����ɢ��Ҳ��Ⱦ���ӹ����ĵ�һ�����ϡ�
��ľ֬��suberin����Ҫ��һЩC16 �� C18�Ļ�������е���ˮ�ԣ��ڲ������Ÿ���ˮ�����ʡ����ޣ�green cotton���Ĵ����������ж��20����������ľ֬����ά�طָ���ͬ�IJ㡣
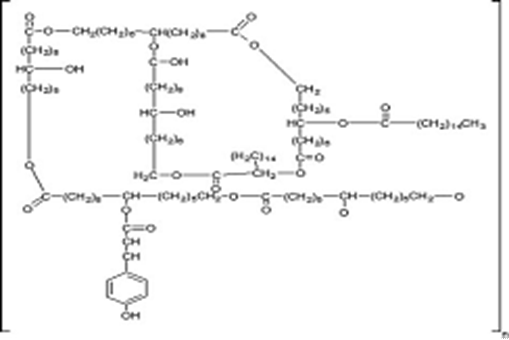
�����أ�cutin����һЩC16 �� C18�����ǻ��ͻ������ĸ�֬���ᣬͨ����������ɵ���ˮ�Ծ��������ʣ���ṹͨʽ�磨1��ʽ��ʾ������C16 �� C18����ɷ����ࣺһ�����16-�ǻ�������9,6-��10,6-���ǻ�������һ��ĵ���Ϊ18-�ǻ������9,10-������-18-�ǻ�Ӳ֬���9,10,18-���ǻ�Ӳ֬���Ρ�
|
|
|
��1�� |
����ά������������2��ʾ��
��2 ����ά��������ϸ���
|
��� |
��Ҫ��� |
����% |
���������ϸ��� |
||||
|
��� |
����ʽ |
������ |
�۵�(��) |
��ע |
|||
|
������������ 52-62% |
��֬���� C23toC34 |
40-52 |
��-��ʮ�� |
C30H61OH |
438.8 |
87 |
������ |
|
���ѷ�(��ɫ����) |
C30H30O8 |
518.5 |
184-214 |
������ |
|||
|
��ú��(1-��ʮ���鴼) |
C28H58O |
410.8 |
83 |
������ |
|||
|
��ʮ���鴼 |
C28H58O |
410.7 |
83 |
���� |
|||
|
������ |
C28-C30 |
- |
- |
- |
|||
|
����(�ͼ�����֬�������ɵ���) |
|||||||
|
���ͼ����� ��֬���� |
7-13 |
��ʮ��(̼)�� |
C27H56 |
|
|
|
|
|
��ʮ(̼)�� |
C30H60 |
|
|
|
|||
|
��ʮһ(̼)�� |
C31H64 |
|
|
|
|||
|
��ʮ��(̼)�� |
C32H66 |
|
|
|
|||
|
ֲ��̴� ��,�̴� ����,������ �Ͷ���ϩ�� |
3-8 |
��-���� |
C20H50 |
414.7 |
|
|
|
|
��-���� |
C20H50 |
414.7 |
|
|
|||
|
���� ���� |
C35H60O6 |
|
|
|
|||
|
����������֬�� |
C30H50O |
|
|
|
|||
|
����������37-47% |
(��ż��̼ԭ �ӵ�)����֬ ����������� |
23-47 |
����ʮ����(ľ������) |
C21H43O20 |
368.6 |
84 |
������ |
|
��-ʮ���� |
C15H30O2 |
242.0 |
69.6 |
���� |
|||
|
ʮ����(�����) |
C16H32O2 |
256.4 |
64 |
������ |
|||
|
˳ʽ-12-�ǻ�ʮ��̼ϩ-9-�� |
C18H34O2 |
282.5 |
04 |
���� |
|||
|
ʮ����(Ӳ����) |
C18H36O2 |
284.5 |
70 |
������ |
|||
|
��-��ʮ����(ɽ��) |
C22H44O2 |
340.6 |
80 |
���� |
|||
|
��-ʮ���� |
C15H30O2 |
242.0 |
69.4 |
���� |
|||
|
ʮ����(�����)ԭ���ظ� |
C16H32O2 |
256.4 |
64 |
������ |
|||
|
|
��֬�� |
�� |
��ɫ��֬������ |
|
|
|
|
���ģ�����ά�ij������ڼ������
������������ڲ���dz������ڣ����ȴ�ԼΪ0.5 ��m������ά�ش���ӿɼ��Եĵڶ������ϡ���������Ҫ�ǹ���������ά�غ��ǵ��ף�������չ������extensin���ȷ���ά��������, �ڳ������������������ε���ά��ԭ�ˣ�����ά�ط��Ӿۺ϶�Dp��2000�����ڴ������ڵģ�Dp��5000���ͣ���������ذ�������������ά�������ʵĻ����С�
����ά�������ڵĽṹ����ϸ���,��ͼ3�ͱ�3��ʾ��
|
|
|
ͼ3 ����ά�������ڵĽṹʾ�� |
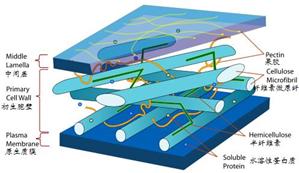
��3 �����������ά�ij��������е���ϸ���
|
|
����ά���������и���ɵĸſ����£�
1������ά��������ά��������ϸ������һ���Ӿ۶�������ľ�ǡ��������ǡ���¶�ǺͰ����ǵȼ��ֲ�ͬ���͵���̼�Ǻ���̼�ǹ��ɵ����Ƕ���壬������ľ����Ϊ��������������ά��ԭ��(microfibril)�ı��棬��������γɽ�����
2���ǵ��ף�������չ���ף��ڳ���������Լռ15%�����ǵ����������Ϻ���������չ�ĵ;��Dz����ĸ�״�����Ҳ����Ϊ�ǻ���������,���е;���Ҫռ65%��
3�������ʣ�pectin����һ��ֲ���Ը߷����Ӿ۶��ǵ��ܳƣ������ԣ�������ά�������ں;��Ʋ�ĵ���Ҫ��֣�������ά����������,����Ϊһ��ˮ���hydrate agent����������ˮ��Һ�������������������ά���˶Ⱥͽṹ����ҪӰ�졣
�������ڹ�����
��һ�����
�����ʵĻ�ѧ���ʮ�ָ��ӡ��ڻ�ѧ�ṹ�ϣ������ǹ����ᣨpectic acid������ѧ���۰�����ȩ�ᣨpolygalacturonic acid������δ�����ľ۰�����ȩ�ᣨLM pectin����������е��Ȼ������������ᣬҲ������Na+��K+ ��Ca2+�������Σ��������д��Ȼ������ľ۰�����ȩ������-1��4���ռ���϶��ɵ��Ӷ��ǣ�heteropolysaccharide���������ӣ�HMpectin��������Ȼ��Ĺ����У���Լ80%�۰�����ȩ����Ȼ��������������ָ������Ĺ�����Ϊ��pectinates��������������С��5%�����֮Ϊ�������Σ�pectates����
������ά�������ڵķ���ά������У�����������Ҫ����ӵġ��ڳ������ڵ���㣬��Ҫ�Ǹ�����������HMpectins��������Ϊһ�����ۼ�������������ά����־ۼ���һ�𡣳��������еĹ����������и��ж��ǵ;�������������������������еIJ�������Զ�ȵ������Ĺ�����������еĴ���Щ�����������ϳ��IJ�����ʹϸ�����ɳڣ���������ά����ʱϸ���ڵ����š��������������ȶ�����һ����ʪ�Ⱥ��¶��£����������룻�ڽϵ��¶ȵļ��Խ����У��������������еļ�Ҳ�������ͨ��������Ӧ�����롣ijЩ�ܽ���������ѽ�ø������-������Ӧֻ�����ڷ���������������л����ڵ��ռ��ϣ��ҹ����ļ������̶�Խ�ߣ�������Ӧ�Ľ�������ҲԽ�졣�ھ��Ʋ��У���Ҫ��δ�����������Ⱥܵ͵Ĺ�����������ˮ�����Թ������������и���ɣ���Ca2+�Ƚ����������к�ǿ�����������Ӷ�ʹ���ڵ����Թ������������γɽ�����ʹ���ԵĹ�������ӽ����һ����ͼ4��ʾ��
|
|
|
|
ͼ4 ���Թ�����������Ľ���ʾ��ͼ |
|
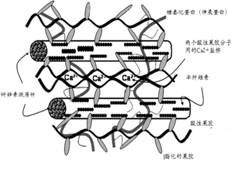
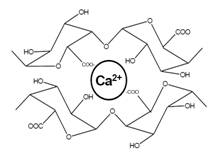
Ϊ�����˽��йع����༸����������Ӣ��������������£�
��1�� Protopectin��ԭ����������������ֲ����֯�в�����ˮ�Ĺ����ʣ�����������ˮ���£�ԭ�������ɹ���(Polymethyl galacturonate)������ᣨPectic acid����
��2�� Pectic acid�������ᣩ���ǿ����Եİ�����ȩ��ۺ����ֻ���пɺ�������negligible amount��-OCH3���ţ���������������γ�Ϊ�������Σ�pectates����
��3�� Pectinic acids����������;�������ᣩ���۰�����ȩ��������к���>0��<75%�������ľ۰�����ȩ�ᣬ��������γ�Ϊ���������Σ�pectinates��
��4�� Pectin������Polymethyl galacturonate����������ȩ��ۺ�������75%���Ȼ�����������
�������ṹ
�ڹ������������ϻ��������ǡ��������ǡ������ǡ�ľ�ǡ������ǡ��۲��ǵ���ɵIJ�������������ͷ��������˰ѹ�������������������ṹ��û�в�����һ��������ȩ����� (Homogalacturonan ���HG)���ƹ⻬����smooth region�����в�ͬ�����ɵ������ǰ�����ȩ�����I (Rhamnogalacturonan I-���RGI)�������ǰ�����ȩ�����II( Rhamnogalacturonan II���RGII)��������ά�Ĺ�������ӵ������У�60%�İ�����ȩ��л��Ǽ��������ǵĻ�ѧ�ṹ�ֱ���ͼ5��ʾ��
|
|
|
ͼ5 ����ά�Ĺ�������������ṹʾ�� |
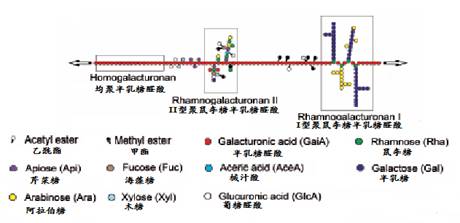
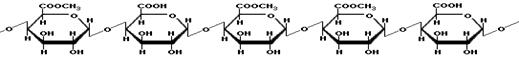
��1��ͬ�Ͱ�����ȩ�����(HG)
HG��D-������ȩ������-1��4���ռ����ӳ���״�ľ��۶��ǣ��䲿��-COOH��������/������������������δ�����İ�����ȩ�Ჿ��Ҳ�������ơ��ء����γ��Ρ��Ǵ����������ƽ̹����������ṹ�磨2��ʽ��ʾ��
|
|
|
|
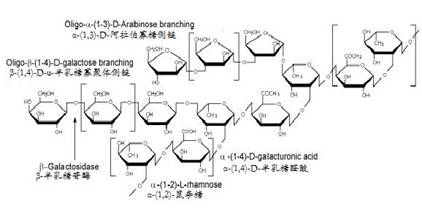
��2�������ǰ�����ȩ����� I��RGI ��
RG I����-(1,4) ������ȩ�����-(1,2)��������ɵ�����������-(1,3)-���������Ǻ���-(1,4)-�����ǹ���������������ṹ�磨3��ʽ��ʾ��
|
|
|
|
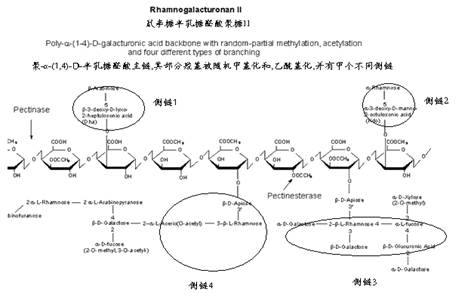
��3�������ǰ�����ȩ�����
II��RGII��
�����ǰ�����ȩ����� II
RGII����-(1,4) ���۰�����ȩ���������ж��11�ֵ�����ɵ��ĸ�����.��ṹ�磨4��ʽ��ʾ��
��-��-��1,4��-D-������ȩ���������䲿���Ȼ�������������������������м�����ͬ����
|
|
|
��4�� |
������֯Ʒø����������Ҫ��֡�������ø����ø
���˽�����ά�Ľṹ��ɺ�ӡȾ�����߾Ϳ��Բ�����֯Ʒ��ø���������У�ѡ����Щ����ø��ȥ������ά������㣨cuticle�������ʺͳ������еĹ����Ȱ�����Խ������ά�����ͳ������ڶԴ�����������ά�ش���ӵ�����������������ʹ��ά�ط������ϵ��ǻ���¶��������ˮ�ȼ��Է��Ӷ����ǻ��ܼ�������ˮ����ʹ�������ںͷ������������Ͽ���������ˮ����ά���ٽ�Ⱦ�Ϻ��й���������������ά�ڵġ���������Ϊ������ӡ����Ⱦɫ�ͺ����������У�Ⱦ�Ϻ��ֻ�ѧҩ������ά�ش���ӵ����ýӴ����������
Agrawal�������֯��ø�����ļӹ�·�ߣ���ͼ6��ʾ��
|
|
|
ͼ6 ��֯��ø�����ļӹ�·�� |
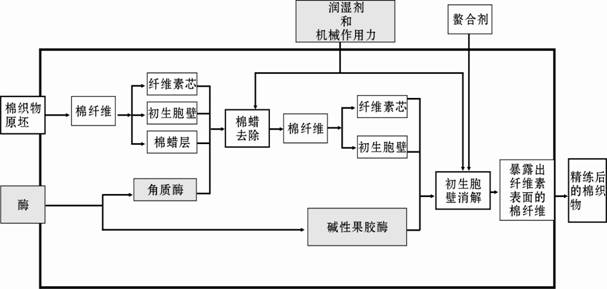
��һ������ø(cutinase
EC 3.1.1.74)
��ø������������У�����Ҫ�������ά�ɼ��Եĵ�һ����������������㡣���������Ľṹ����ɣ�ɸѡ�ܴ�֬��������ˮ�ⷴӦ��ø����ø�����У�����ø��ˮ����ø��EC 3.1.1,X�������п�ѡ�õ���������ø(carboxyl esterase EC 3.1.1.1)��֬��ø(lipase EC 3.1.1.3) �� ����ø(cutinase EC 3.1.1.74) ����������ø�У�������ø�Ծۼ�����ɢ��֬�����������õ�ø�ٻ��ԣ����Ծۼ����ܵĽ��ʲ����롣֬��ø�ͽ���ø�Ծۼ����ܵĽ��ʶ��иߵ�ø�ٻ��ԣ���֬��ø�Ƿ�ˮ���Եģ���ø�ٷ�Ӧ�У���Ҫ����һ��֬�ʺ�ˮ����Ļ�ܣ���ȥ����������ڴ���������������ʹ֬��ø�Ļ������������Ӵ�������ø�ٷ�Ӧ��������ø��Ҫ��ͬʱ��ˮ��ʱ������ҪCa2+����������������ø���á��ɴ˿ɼ�������ø�ǽ������ȥ������ά������ø��
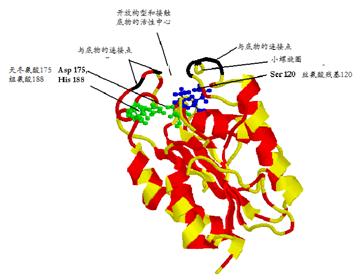
����Ȼ���У�����ø������һЩֲ�ﲡԭ�������ϸ���У��縯Ƥ��������Fusarium solani Pisi��������ϸ����Thermobifida fusca�������ڽṹ�ϣ�����ø��cutinase�����ɵ��͵�˿����-�鰱��-�춬��������Ԫ��Ser, His, Asp triad����ɵ�ˮ��ø����������ø�ٻ���������˿����л�����ʾ��ͼ��ͼ7��ʾ
|
|
|
ͼ7 ����ø�Ľṹʾ��ͼ |
��1������
����ø���Խ��ʣ���Ҫ�Ǹ�֬����������ˮΪ����ƻ�ֲ�����Ƥ���Ӿ۾������ӵ�������ʹ��ˮ��Ϊ�����С���ӹѾ��塣�����ľ������ԣ�����ø�ܴ�����ά������н������֬��������ˮ�ⷴӦ���ƻ�����ά���Ƥ�в����Ը������������������ͽ��������ˮ�Ծ����������������ά�ɼ��Եĵ�һ�����������������ھ���ʱ����ø�Թ����Ŀɼ��ԣ��Ӷ��ﵽ�����õľ���Ч��������Agrawal�����������ø����ø�����ǽ�����ù���ø��������֯����ʪ�Բ��ѵ���Ҫ���ԡ�
2002��Degani �����״α����÷����ѹҺ��ɫ����(reverse phaseHPLC)������ɫ��-�����ǣ�GC-MS�������ٲ���������ˮ���ͷŵ�C16��C18֬����ķ������о��üٵ�������ϸ����Pseudomonas mandocino�������Ľ���ø������֯����֤ʵ����øȷ��������ȥ����ø�ٷ�Ӧ���ԣ���ʱ�������10-20��Сʱ��������ʵ��Ӧ�á�
2007��Agrawal�����ø�Ƥ��������Fusarium solani pisi�������Ľ���ø��100 U/g֯���pH8��50 mM Tris-HCl�����Ǽ���������-���Ỻ��Һ)����Һ�У�30���½��账��15-30���ӣ�ͨ���ⶨ֯��ṹ�Ӵ���*��ɨ����������۲��������������ý���ø����Ч��ȥ����֯���е����ʣ���ȥ��Ч��ԶԶ����Degani�������ģ��������鴦���Ľӽ���
ͼ8Ϊ��֯���ý���ø����������-������ȡ��֯��ṹ�Ӵ����ȽϽ����
|
|
|
ͼ8 ��֯�ᆳ����ø��������������ȡЧ���Ƚ� ��A������ø100
U /g_֯���pH 8��50 mM, Tris-HCl��30�洦��30���ӣ� ��B������-���� ��75oC��30������ȡ��֯� ��C���հ� |
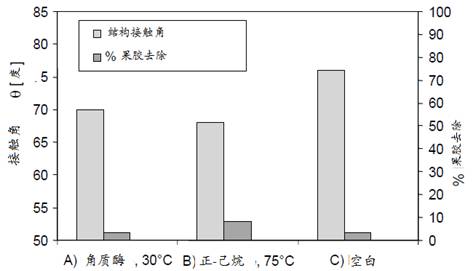
��ͼ8�ɼ����ý���ø���������֯���ṹ�Ӵ���Ϊ70����������-������ȡ��68���dz��ӽ�������ȥ���ʶ��ܵ͡�
��*������������֯�о�Ժ���Զ���϶�ǣ�TRI auto-porosimeter���ⶨ�ṹ�Ӵ��ǣ�֯���п���Ч�뾶Reff=2��cos��/��P��ʽ������������P�ֱ�Ϊ�����������Ӵ��ǺͿ�����ѹ������ŵ��Dz�õĽӴ�����֯���ɴ���ں�ɴ���϶��С���ֲ���ϵ����
��ͼɨ��羵ͼ9�ɼ����հ�������C������ά�����й⻬�������㡣������ø�����ģ�A��������-������ȡ��B��������ά����⻬�������������ʧ����¶�������ֲڵij������ڡ�
|
|
|
ͼ9 ��ͬ��������ά����ĵ羵ɨ�� |

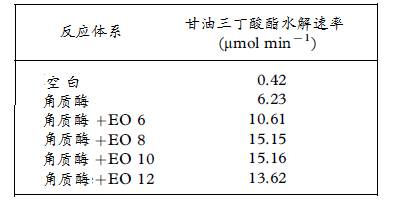
Ϊ����߾������ȶ�֯�����ʪ�ԣ��ھ���Һ�ж����б�����Լ����������ӱ�����Լ��ή�ͽ���ø���ԣ�����ʹ�á�Agrawal�ò�ͬ����ϩ��EO�����ķ����ӱ�����Լ������ø���ã��Ը�������������tributyrin ��Ϊ����ⶨ��ˮ�����ʺ�ˮ��ٷ��ʡ����Դ��䷽�Ĺ���Һ������֯��о���ͬ����ϩ��EO�����Խ���ø���Ե�Ӱ�켰�侫��Ч����������4��ͼ10��ʾ��
��4 ��ͬ����ϩ���Խ���øˮ�����ʵ�Ӱ��
|
|
|
|
|
|
|
ͼ10 ��ͬ����ϩ���ķ����ӱ�����Լ��Խ���øˮ�����ʵ�Ӱ�� |
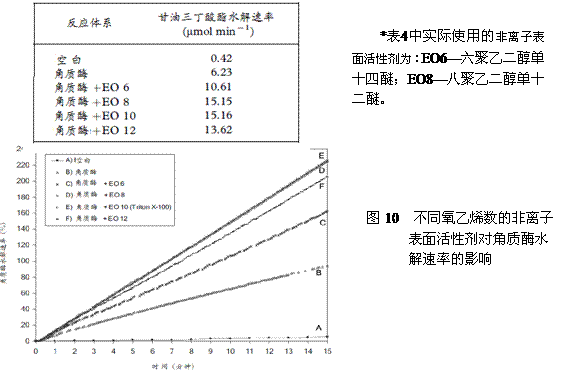
��Agrawal��֯��ṹ�Ӵ�����������ͬ����ϩ��EO���������ӱ�����Լ�����ʪ������ָ����E06��E08��E010��Triton X-10����E012Ϊ�ã����Բ�ͬ����ø������֯��ĽӴ��ǽ��бȽϣ�������ͼ11��ʾ��
|
|
|
ͼ11 ��ͬ����ø�����ĽӴ��� A�� ����ø����100U/g��30�棬30����PH=8��Tris-Hd���壩 B�� ����������ø����ʱ����Triton
X-100 1g/L C�� ����ø�������پ�������75����ȡ30���� |
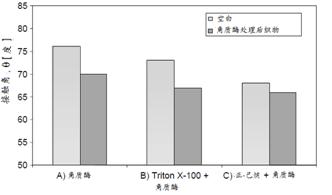
��2������ø�Ļ��Բⶨ�������
���ݹ��������ϣ�����ø�Ļ��Բⶨ������Ҫ���Զ���������������p-nitrophenylbutyrate -PNPB)Ϊ����ķֹ��ȷ����Ը�������������tributyrin
�������pH-stat����
1���Զ���������������p-nitrophenylbutyrate -PNPB)Ϊ����÷ֹ����������ⶨ�ڲ���405 nm������ȣ��ⶨ����øˮ��������ͷŲ���-���������ӵ�����������ø�Ļ������������£��������ֹ��ȷ���20 ���½���. ��Ӧ���Ϊ1 mL������20 ��LøҺ��980 ��L��50 mmol/L����������������pNPB����50 mmol/L������������ơ�20 mmol/L Tris�CHCl����Һ��pHֵ8.0������405 nm������¼�������ӵ���������. ø��壺��20 �棬ÿ���Ӵ���������������ˮ������1 ��mol�������ӵ�ø����Ϊһ��ø������λ.
2��2007��Agrawal���˱�������Vertommen���� ��2005����ܵķ������Ը�������������tributyrin ��Ϊ�����pH-stat*�����ϣ�30oC��20����������0.01M��NaOH����Һ�ζ�����������ø�Ļ��ԡ���1����ø�ٸ�����������ˮ����ͷ�1 mmol ֬�����ø����Ϊ1��λ(U)���˷���Ҳ���ڲⶨ����ø����֯���н������Լ��ڸ��ַ����ӱ�����Լ�����ʱ��ˮ����ԣ��䷽���������£���pH-stat*�����ϣ�ȡ0.5����֯�����ں��н���ø��pH9.5��2mmol��ɰ����Һ�У���30oC����15���ӣ�����pH9.5������֯���϶���C16֬�����pKֵ������ˮ���ͳ��������������ɣ�����pH-stat*�����ϣ�30oC��20����������0.01M��NaOH����Һ�ζ�C16-18֬���ᣬ�о�����ø����֯�����ʵ�ˮ����ԡ�
��*pH-Stat����������������ע�����ᣬ�кͷ�Ӧ�������ͷŵ�����Ա��ַ�Ӧ��ϵpH�㶨���Ӷ��������ü����������ⶨ��ѧ��Ӧ����ѧ��һ�ַ�������
��3������ø������Ӧ�ò���
ͨ��pH���¶ȡ�Ũ�ȡ��ʹ���ʱ������飬�����������Ӧ�ò�����
����pH�������ø���������������������������ֵ���������ø��ø�ٷ�Ӧ�������飬��õ�����pH�ֱ�Ϊ9-10��8.5����������pHΪ8-9��
�����¶ȣ�30oC������40oC, ����øʧ�
����Ũ�ȣ�����øŨ����100U/gʱ���䴦��֯��Ľṹ�Ӵ�����С��70o��
��Ѵ���ʱ�䣺15���ӡ�
�ڴ���ԡ�У�������ʵķ����ӱ�����Լ�����ѡ���������ӱ�����Լ����ã���������߽���ø�Ĵ���Ч����
����������ø(Pectinases)
��1�����������
����ø��һ���Թ���Ϊ���ͨ��ø��ˮ����ѽⷴӦ��ʹ����ȥ�������ͽ��⣬���ɰ�����ȩ��;��������ȩ��������ȩ�ᣨ�ף����Ķ���ָ�����ø���ܳơ�
�ù���ø����������ȥ������ά�������ں;��Ʋ��еĹ����ʣ�Ҳ��ʹ���а���ά�غ��ǵ�����������ά�������ʴӹ������н��ѳ�������Ч�Ľ��Ⱦ���ӹ��ĵڶ��������������ﵽ����Ч����
�ݱ���,������ʮ���������ܲ�������ø,�����о���Ӧ�ý϶����ù������ø,�����ϸ������ø������ù������ø����������ø,��ϸ������ø����Ҫ�Ǽ��Թ���ø��
��Ȼ���У�����ø�㷺������ֲ���Լ������ϸ����������(ͨ������ϸ�����ܺϳ�����ø),��Ҫ��ʹ�����������ѳ����Ĺ�����ø��pectin esterases����ʹ����������1,4�ռ�ˮ��ľ۰�����ȩ�ᣨˮ�⣩ø��polygalacturonases���PG����ͨ��������Ӧʹ����������1,4�ռ��ѽ�ľ۰�����ȩ���ѽ�ø(Polygalacturonate lyse���PGL)��
��EC�����У�������ø�;۰�����ȩ��øͬ��ˮ��ø�࣬����ˮ�����ͷ�Ӧ��λ��ͬ�����Ƿֱ����Ϊ��������ø��EC 3.1.1.11�������о۰�����ȩ��ø(endo PG��EC 3.2.1.15)�����о۰�����ȩ��ø��exo PG��EC 3.2.1.67)��
������ø��pectin esterases����
EC3.ˮ��ø��Hydrolases. ��
3. 1. �������������Acting on ester bonds ��
3. 1. 1. ������ˮ��ø��Carboxylic ester hydrolases ��
3.1.1.11 ��������ˮ�⣩ø��Pectinesterase ��
�۰�����ȩ�ᣨˮ�⣩ø��polygalacturonases����
EC 3. ˮ��ø��Hydrolases��
3. 2. �ǻ���ø��Glycosylases��
3. 2. 1.-����ø��Glycosidases������ø�ٷ�Ӧʹ�ǻ������� O- �� S-��ˮ��
3.2.1.15 ���о۰�����ȩ��ø��Polygalacturonase ��
3.2.1.67 ���о۰�����ȩ��ø��G alacturan 1,4-alpha-galacturonidase ��
3.2.1.82 ���о۰�����ȩ��ø��exo-poly-alpha-galacturonosidase�����ڷǻ�ԭ��ˮ������������ȩ����ǡ�
��EC�����У������ѽ�ø���ѽ�ø�ࣨlyase����EC����ΪEC.4.2.2.X, ����ˮ�ⷴӦ�IJ�λ��ͬ,�ַ�Ϊ: ���о۰�����ȩ���ѽ�ø(endo PGL��EC.4.2.2.2)�����о۰�����ȩ���ѽ�ø(exo PGL��EC.4.2.2.9) ���߾�Ϊpectate lyase���ۼ�������ȩ���ѽ�ø(�ֳ�pectin lyase),������ֻ�������;ۼ�������ȩ���ѽ�ø(Endo PMGL��EC.4.2.2.10��)���������ͣ�Exo PMGL��Ŀǰδ��������
�����ѽ�ø(pectate lyase��pectin lyase)��
EC
4.-Lyase �ѽ�ø-ͨ����Ӧ����ˮ���������Ӧ�⣩������C-C, C-O, C-N ����ø
4.2.-������ C-O�����ѽ�ø��Carbon-Oxygen Lyases��
4.2.2.����Ϊ����ǣ�Acting on Polysaccharides��
4.2.2.2
���о۰�����ȩ���ѽ�ø��pectate
lyase��
4.2.2.9���о۰�����ȩ���ѽ�ø��pectate disaccharide-lyase�����ֳƹ������ζ����ѽ�ø
4.2.2.10
���оۼ�������ȩ���ѽ�ø��pectin
lyase��
����ø�������5��ʾ��
��5 ����ø����
|
|
|
|
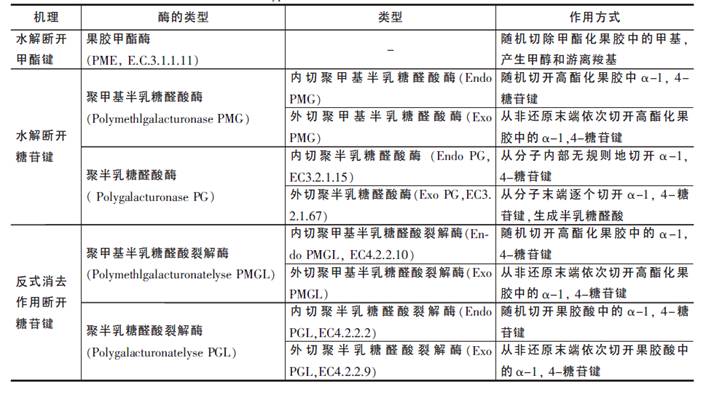
��2������ø��ø�ٷ�Ӧ
1��������ø��EC3.1.1.11����������ˮ��ø����ʹ�߶������Ĺ����ϵ�����ˮ����ȥ�������ɾ۰�����ȩ�ᣨ�Σ��ͼ״���
������ø�Ļ�ѧ��Ӧ�磨5��ʽ��ʾ��
|
|
|
��5�� |
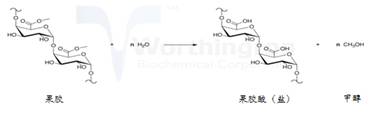
���ڹ�����øֻȥ�������е������ţ��������������ӡ���˲�������Ϊ�ľ�������
2���۰�����ȩ��ø(polygalacturonidases��� PG)
�۰�����ȩ��ø��PG)��ø�پ۰�����ȩ�����-1,4�ռ�ˮ�ⷴӦ��һ�ֹ���ø������ˮ�����õIJ�λ��ͬ���ɷ�Ϊ���о۰�����ȩ��ø(endo- PG�� E.C.3.2.1.15)�����о۰�����ȩ��ø(exo- PG�� E.C.3.2.1.67)������ø�ֿɰ�øԴ��ͬ���ֳ��������ͣ�һ����������о۰�����ȩ��ø���������ղ����ǰ�����ȩ�ᵥ�壻��һ����ϸ�����о۰�����ȩ��ø��EC 3.2.1.82�����������ղ����ǰ�����ȩ��Ķ����塣
�۰�����ȩ��ø���ﻯ���������ܿ��ԴӲ�ͬ����Դ������۰�����ȩ��ø�������ǵ��ﻯ���������ܺ�����ģʽ�����ԵIJ��졣���д����PGø���������pH�����Է�Χ��3.5-5.5���������˵��¶ȷ�ΧΪ30-50�档Ҳ������,�������ѿ�߸˾������꣨ BacillusLicheniformis����������������Fusarium oxysporum�������PGø����������������ڼ��Է�Χ��pH11���ͽϸߵ��¶ȣ�69�棩��
���о۰�����ȩ��ø��ѧ��Ӧ�磨6��ʽ��ʾ��
|
|
|
��6�� |
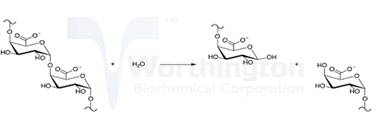
���о۰�����ȩ��ø�ķ�Ӧ��λ�ڹ������������ڲ������ɷ������϶̵Ĺ����;���,�������з�Ӧ���������ۺ϶Ȳ��Ͻ��ͣ����ɰ�����ȩ�ᵥ�塣
���о۰�����ȩ��ø��ѧ��Ӧ��
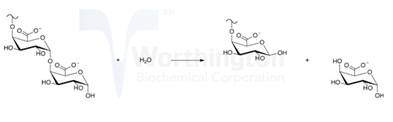
��������ڵ����о۰�����ȩ��ø���䷴Ӧ��λ�����ڹ����������Ķ˲������ɰ�����ȩ�ᵥ�壬�磨7��ʽ��ʾ��
|
|
|
��7�� |
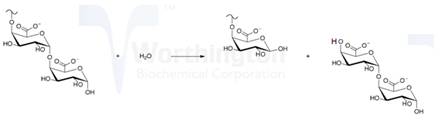
��Ҳ��ijЩϸ�����ڵ����о۰�����ȩ��ø���䷴Ӧ��λҲ�ڹ����������Ķ˲��������ɵIJ����ǰ�����ȩ������塣��Ӧ�磨8��ʽ��ʾ��
|
|
|
��8�� |
��3�������ѽ�ø(pectin
lyases��PL)��ø�ٷ�Ӧ
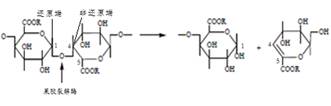
�����ѽ�ø(�ֳƷ�ʽ��ȥø)��������ȥ��Ӧ����-elimination���������ѽ��һ�ֹ���ø���ѽⷴӦ�����ڹ������������������ڰ�����ȩ��л���ǻ�ԭ��һ��л���C-4���ռ�������Ӧʹ��л���C-5��ʧȥһ��Hԭ�ӣ�������ԭ�˲л���C-1����O����γ�-OH,ͬʱ�ڷǻ�ԭ�˲л���C-4��C-5�����һ�������ͼ������ɲ����͵���4,5-D������ȩ���磨9��ʽ��ʾ
|
|
|
��9�� |
���ݹ����������Ⱥͷ�Ӧ�IJ�λ�������ѽ�ø�ֿɷ�Ϊ��
1�������ᣨ�Σ��ѽ�øPectate Lyases
��Ϊ�����ᣨ�Σ��ѽ�ø��PGL�������ȣ�preferential������Ϊδ�����������Ⱥܵ͵ĵĹ�������Ӣ����ΪPectate lyase����ͨ����ʽ������Ӧʹδ�����������Ⱥܵ͵Ĺ����۰�����ȩ�ᣨpolygalacturonic acid�������ռ����ѣ����ɲ����͵���4,5-D������ȩ���Ρ������䷴Ӧ��λ��ͬ�����������ѽ�ø�ɷ�Ϊ����1�����о۰�����ȩ���ѽ�ø(endo PGL��EC.4.2.2.2)���䷴Ӧ��λ����ڷ������м䣻��2�����о۰�����ȩ���ѽ�ø(exo PGL��EC.4.2.2.9) , �䷴Ӧ��λ�ڷ������Ķ˲�������ϸ�������о۰�����ȩ���ѽ�ø�������ѽ����Ϊ�����壬�������˳���Ϊ�������ζ����ѽ�ø��Pectate disaccharide-lyase����ֵ��ע����ǣ������ᣨ�Σ��ѽ�ø��ø�ٷ�Ӧ������Ca2+������EDTA���Ҷ��������ᣩ�Ƚ������ϼ�����ø�ٷ�Ӧ�к�ǿ���������á�
2�������ѽ�øPectin Lyases
��Ϊ�����ѽ�ø�����ȵ���Ϊ�������ȵĹ���������Ӣ����ΪPectin lyase����ͨ����ʽ������Ӧ�������ʹ�������ȵĹ��������ۼ�������ȩ�ᣨpolymethylgalacturonic acid�������������ռ����ѣ����ɲ����͵���4,5-D��������ȩ��;���ݳƣ�Ŀǰ�������Ĺ����ѽ�ø���������й����ѽ�ø��endo PL,EC.4.2.2.10��, ��ø�ٷ�Ӧ��һ����ҪCa2+�����ɱ�Ca2+�����������Ӽ�������Van Alebeek���о��������ɺ���ù��Aspergillus niger���Ƶõ������ѽ�ø����ø�ٷ�Ӧ����Ϊ�����Ͱ�����ȩ��Ķ��ۡ����ۺ��ľ����δ��������Ͱ�����ȩ��ĵ��塣
�������ѽ�ø��CAZy�����У�����Ϊ��һ�壬������ѽ�ø��family 1�� polysaccharideslyase����������Ƶõ��ѽ�ø������ѻ��������Ժ����Խ��ʣ�����ϸ�������Ƶõ��ѽ�ø������ѻ����ڼ��Խ����С�
�����о���Ϊ����-������Ӧֻ�����ڹ�������������������Ȼ����ڵ����ռ��ϣ����Թ����ļ������̶�Խ�ߣ��併���ٶ�ҲԽ�졣
���������������ѽ�ø����pH�ڼ��Է�Χ���������˰ѹ����ѽ�ø��Ϊ���Թ���ø��Alkaline pectinases������������pH�����Է�Χ�ľ۰�����ȩ��ø��Ϊ���Թ���ø��Acidic pectinases��������ʵ��ȴ���������⣬�������ѿ�߸˾������꣨BacillusLicheniformis���� ������������Fusarium oxysporum��������۰�����ȩ��ø��������pH=11��
��4������ø������ø�ıȻ�������ⶨ
ø�����DZ���ø�ٷ�Ӧ�Ĵ�����������ø������λ��U�������ǣ����ض�������25�棬����Ϊ�����������£���1��������ת��1��mol�����ø��������ת��������1��mol���йػ��ŵ�ø�������ڰ��˶����ʾ������Ҫ��С�����U��ʾ�������㡣ø��Ӧ�������ã�1������ת��1nmol������йػ��ŵ�ø��������ø�����ĵ�λ��
ø�ıȻ�����specific activity������ָÿ���ˣ���������������������ij��ø�Ĵ�������U/mg��������������ø���ȵ�ָ�ꡣ��������øѧ�о��о���ʹ�õĻ������ݡ�
����ø�����ⶨ���������Ȳⶨ��ͬʱ��ø�ٷ�Ӧʱ���ɵIJ���Բ���Ũ��Ϊ�����ꡢʱ��Ϊ�����꣬����ø��Ӧ��������ͼ���ڷ�Ӧ��ʼ��ijһ�϶�ʱ����ڣ����߳�ֱ�ߣ���ʾ����������ٶ���ʱ������ȣ���б�ʴ���ø�ٷ�Ӧ�ij��ٶȡ����ŷ�Ӧʱ���ӳ�������Ũ�Ƚ��ͺͲ���Ũ�����ߣ������淴Ӧ��ǿ����Ӧ�ٶ��½�����ʱ�������ߵ�����б����С��ֱ����������ƽ̹����ʱ��ʾ�ټ����ӳ�ʱ�䣬��Ӧ�ٶȼ������䡣���Բⶨø�����ķ�Ӧʱ��Ӧѡ�������߳�ֱ�ߵ���һʱ����ڡ�
�۰�����ȩ��ø(EC3.1.1.15)�Ļ������Է���
������ʳƷ��ҵ�ù���ճ���½������Ծ۰�����ȩ��ø(EC 3.1.1.15)�Ļ����⣬�ڻ�ѧ�������棬��Ҫ�Ǹ��ݾ۰�����ȩ��ø�Թ�����ø��ˮ�����--D-������ȩ����л�ԭ�ԣ����ø��ֻ�ԭ�ǵIJⶨ�����������������
������λ��U���Ķ��壺��37���pH=5������,1���ӴӾ۰�����ȩ��ֽ��ͷ�1��molD-������ȩ���ø���������Ǽ��ֲ��Է�����飺
1�� DNS (3,5��������ˮ����)������Ӧ�ý϶�ķ���֮һ��
ԭ������NaOH�ͱ����������£�3,5��������ˮ���ᣨDNS����۰�����ȩ���ø�����--D-������ȩ�ᣨ��ԭ�ǣ����Ⱥ���ԭ���ɰ���������ڹ�����NaOH������Һ�д˻�����ʽۺ�ɫ����540nm��������������գ���һ����Ũ�ȷ�Χ�ڣ���ԭ�ǵ���������ȳ����Թ�ϵ���÷ֹ��ȼ�������D-������ȩ������ߣ�Ȼ��ⶨø�ٷ�Ӧ����Ʒ�е�D-������ȩ�Ậ�����䷴Ӧ�磨10��ʽ��ʾ��
|
|
|
��10�� |
2��I2-Na2S2O3����-��������ƣ��������ڹ���ø�Ծ۰�����ȩ���ø�����--D-������ȩ����л�ԭ����ȩ��,���ô��ǵ��ᷨ �����ⶨ���Ե���Ϊָʾ������Na2S2O3�ζ�������I2�����л�������ҵ�� QB 1502-92������ø������������Sigma-Adrich�ֲᣩ��
3������ͭ�Լ����������Լ�CuSO4�е�Cu2+����л�ԭ����ȩ����ø�����-- D-������ȩ�ᷴӦ����Cu+����������ͭ�Լ�������ɫ������ڲ���Ϊ450nm�������շ壬��һ����Ũ�ȷ�Χ�ڣ����������A�����Թ�ϵ���÷ֹ��ȼƲⶨ��Ʒ�е�-D-������ȩ�Ậ������Worthington �ֲᣩ��
��5�������ѽ�ø�����ⶨ
ԭ�����ڹ����ѽ�ø��ø�ٷ�Ӧ�£��������Ӻ�ˮͨ��������Ӧ������-4,5ϩ���İ�����ȩ�ᵥ���;���ڲ���235nm���в����͵���4,5-D������ȩ���������շ壬�÷ֹ��ȼ������ⶨ5���ӣ����߳����Է�Χ�ڣ��������ѽ�ø��ø�ٷ�Ӧ��������ÿ�����������ֵ����A235nm/���ӣ�Ϊ1����Ϊø�Ļ�����λ��
�ġ�����ø�����ѽ�ø��������Ӧ��
��������ø����ǰ��ֵ��һ����ǣ�1�����۴������ϸ�������ƵõĹ���ø��Һ�����Ǿ��������ᴿ��ø�Ƽ������ǹ���ø������ø�Ļ���Ҳ������ijһøΪ��������������ø�Ļ���塣2����ʹ��ͬһ��ø��ͬһEC��ţ����������������������Ҳ��øԴ�����꣩��ͬ���кܴ���졣3��ø��������Ϊ�����ø�Ƽ��ľ���Ч������Ʒø�Ƽ����Ǽ���ø����Ļ���壬���л������ȶ������黯���ͱ�����Լ��ȡ�4��ø������ø�ıȻ�����ø�Ƽ�Ʒ�ʵ���Ҫָ�꣬���о�����������ø�Ƽ�ʱ��ø�����ⶨ�DZ�������Ŀ֮һ,���Dz��������������Ũ�ȡ��¶ȡ�������ȣ���ͬ,��õ����ݲ���ܴ�����ӡȾ����ʹ��ijһ���ŵĹ���ø�Ƽ�ʱ������Ҫ�˽���������ø�Ƽ������ˮ��ø�����ѽ�ø����ø����������������������ֺ��ȶ��Ե������⣬�ڿ��ܵ�����£���Ӧ��Ӧ���˽���������Է���������������Ļ�����Ӧ��ʵ�����У�ģ��ʵ��Ӧ����������Ч�����ԣ��������ܿ�ѧ����ƹ���Һ��ɺ;�������������
�������Ͻ��ܣ����磨1995�꣩ʹ�ù���ø������ø��֬��ø����ά��ø������ø�����о����ǣ��¹�Schacht�� Rößner���ˡ��˺����ܶ��˵ļ����о�����Li &Hardin, 1998; Hartzell & Hsieh, 1998�� Buchert et al., 2000��Traore & Buschle-Diller, 2000��Galante & Formantici, 2003������Ϊ������ø����ά��ø�ľ���Ч��������֬��ø��֮������ø��
�������������кܶ����Ͻ����ù���ø-����ø������ø-��ά��ø������ø-��ά��ø-����ø����ø-����ά��ø����ϣ��о����ǶԹ���ȥ���ʡ�֯�����ˮ�ԡ��ȡ�֯����������ܱ仯��Ⱦɫ��Ⱦ�Ե����ܵ�Ӱ�졣��Ϊ����ά�������ڵ�����㸻�����ּ������Ĺ����������������ѽ�ø����Novozymes Bioprep 3000L������-������Ӧֻ�����ڹ�������������������л����ڵ����ռ��ϣ�������Ľ���Ч���������Թ���ø�����⣬������ǰ��������ڼ��������½��еģ��ѽ�ø���������ڹ�����ν�,���Լ������ʵ��ѽ�ø��Ϊ��������ע��
�ܶ��о�����Ϊ�����ù���ø�����ѳ�����������ʹ֯�������������ˮ�ԣ�����ø-����ø������ø-��ά��ø����ϡ�����ǰ֯������1000CˮԤ����������ʱ��֯��ʩ�ӻ�е���ͳ������ȷ���������ʹ������֯����кͳ�����ͬ����������ˮЧ����
��һ�����鷽������
Agrawal����νͳһ���鷽��*���Խṹ�Ӵ��Ǻ��ɺ�Ⱦɫ���ⶨ�IJ�������**��Ϊ����Ч����ָ�꣬��A-δ���κδ�����B-����������ȡ��750C��ȡ30���ӣ����ʺ��C-����ͳ�����Ϊ�����������Ƚ��������Թ���ø���������Թ����ѽ�ø��ø����Ч����
��*ͳһ���鷽��������֯��Ϊ280g/m2ȫ��ƽ��֯�������500������ø��pH8��Tris-Hcl������Һ��1000�����ձ���
����3��5��12cm��֯��ڹ涨�����´�����ȡ����������500����900C��ˮ����15���ӣ�ʹøʧ�������ˮ5����ˮϴ���Σ�Ȼ������ƽ����ƽ�����л�������,����24Сʱ����и�����ԡ�ÿ�����鶼�ظ����Ρ���
��**�ɺ�Ⱦɫ���Dzⶨ��������֯���Ϲ�����Բ������ļ�ʵ�÷���������������£�ȡ����Ҫ�ⶨ�����IJ�����һ���������趨��������Ϊ100%���Լ�һ�������IJ������趨��������Ϊ0%����ϳ�ֱ��Ϊ8cm���������˲������밴100ml/g֯��������Ⱦɫ��Һ�С�Ⱦԡ��ɣ��ɺ�0.2g/l+1g/l������Լ����Һ��Silwet
L-77 and Tergitol 15-S-12��������Ⱦɫ15���ӣ�Ⱦɫ��ȡ������������Linitestʵ����Ⱦɫ���У���60���ȥ����ˮϴ��10���ӡ��������ɺ��ò�ɫ���ڲ���Ϊ540nm�����䷴����R������Kubelka Munk*��ʽ�ֱ����K/S����������K/S0�������K/S100��ԭ����ֵ�������й�ʽ���������Ĺ�����Բ�������
![]()
��
��1������������
��A��δ���κδ����������Ľṹ�Ӵ���Ϊ830, �趨�����ȥ����Ϊ0%��
��B������������ȡ��750C��ȡ30���ӣ����ʺ����������ṹ�Ӵ���Ϊ��680���趨������ȥ����Ϊ100%��
��C������ͳ���������Ϊ����ṹ�Ӵ���Ϊ530���趨�����ȥ����Ϊ100%��
�������������ıȽ���ͼ12��ʾ
|
|
|
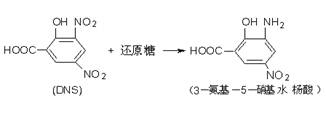
ͼ12 ��ͬ�账��������֯��ṹ�Ӵ���Ӱ�� |
��2��ø������������������øŨ�Ⱦ�Ϊ100 U/g������pH���Թ���øpH5*�����Թ����ѽ�øpH8**��500C,30���ӣ�
��* 50mM�����λ���Һ��**
50mM Tris-HCl����Һ����
���Թ���ø��
�������Թ���ø���Ǵ����Sclerotium rolfsii������С�˾����з��룬���ֱ��������Ǻ���ά�ض��ֲ�ͬ���������Ƶã���
1) ������Ϊ��������PG-glucose�������о۰�����ȩ��ø��EC3.2.1.15��,
2) ��ά��Ϊ������PG-cellulose, �����о۰�����ȩ��ø��EC3.2.1.67��,
3���Ǿ��������Ƶõ�PG-precipitated,Ҳ�����о۰�����ȩ��ø��
������500C��pH5ʱ�İ�˥��Ϊ10Сʱ������pH8ʱֻ��20���ӡ�
���Թ����ѽ�ø
�������Թ����ѽ�ø�ֱ�Ϊ��
1��Novozymes Bioprep 3000L,���ǵ�һ�������ѽ�ø��pectate lyase������Ʒ�Ƽ���������pH8-9��
2��Ϊ����Bioprep 3000L��������ֵ�Ӱ�죬ʹ��ǰ�Ⱦ�����麟����ᴿ��
3���Ӷ�Сѿ���˾���Bacillus pumilus��������ȼ�ϸ�����������й������ѽ�ø��PL EC 4.2.2.2��������pHΪ8.5��Tris-HCl,�����Ǽ���������-���Ỻ����ϵ��������300Cʱ��˥��Ϊ75Сʱ��ø�ٷ�Ӧʱ����ҪCa2+,EDTA��Co2+�Է�Ӧ��ǿ������ЧӦ���������������������ȴ���50%ʱ�����ѽ�ø�Ļ����ή�͡�
��3����������ͼ13��14�ͱ�6��ʾ��
|
|
|
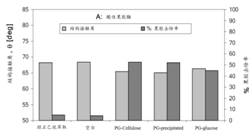
ͼ13 ���Թ���ø�Ĵ���Ч�� |
|
|
|
ͼ14 ���Թ���ø�Ĵ���Ч�� ע��ͼ13��14�С��հס�Ϊ��������ȡ�����֯�����ø�������´����Ŀհ����� |
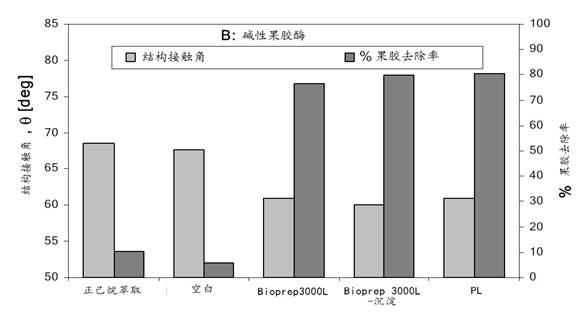
��6 ���ֹ���ø����Ч���Ƚ�
|
|
����ȥ���� |
�ṹ�Ӵ����� |
|
���Թ���ø��PG�� |
45%-52% |
∼66�� |
|
���Թ����ѽ�ø��PL�� |
76%-83% |
∼60�� |
|
δ���κδ��������� |
�趨Ϊ0% |
∼830 |
|
��������ȡ��750C��ȡ30���ӣ������� |
��7% |
∼680(�趨����ȫ��ȥ��) |
|
����ͳ������� |
�趨Ϊ100% |
∼530 |
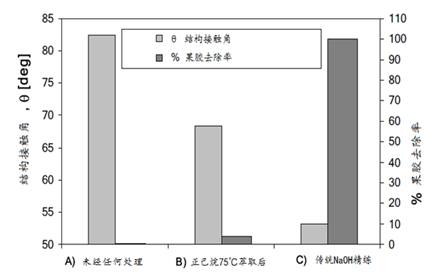
������������1�����Թ����ѽ�ø�Ĺ���ȥ���ʱ����Թ���ø�ߴ�75%���ϣ����߽ṹ�Ӵ��Ƿֱ�Ϊ∼60����∼66����ǰ�߱Ⱥ���С∼6o������С����������ȡ��������∼680����2���������Թ���ø�У� PG-cellulose��PG-precipitated�Ĺ���ȥ���ʸ���PG-glucose�������ߵĽṹ�Ӵ��Ƕ�С����������ȡ��������∼680����3���������Թ����ѽ�ø�ľ���Ч��������ȥ���ʺͽṹ�Ӵ��ǣ��ӽ���
�б����ƣ������������ȸߣ��ή����������ˮ��Ľ����ٶȣ�������pH�ɼӿ���ʽ������Ӧ�ٶȺ������������Ѽ���������������������������ж�����������������Ӽ�����õĶϿ���
�ɴ�Agrawal��Ϊ�����Թ����ѽ�ø�ľ���Ч���������Թ���ø�����Լ��Թ����ѽ�ø��PL������һ����ʵ�顣
����������ȥ���Թ���ø��PL������Ч����Ӱ��
��ǰ����������ά��������ʲ���谭����ø�������ڱ���Ĺ�����������ˣ����ʲ��ȥ�������ڹ���ø���ܵķ��ӡ�Ϊ��֤ʵ����۵㣬���������������飺
A�飺֯���Ⱦ�������ˮԤ����2���ӡ�b��100oC pH8(50mM Tris-HCl)����ҺԤ����2���ӡ�c)��������75oC��ȡ30����,�̺�ֱ��ù���ø��100 U/g PL��50oC 30���ӣ�������
B��:֯���Ⱦ�����ͬ����Ԥ�������̺�ֱ��ù���ø(100 U/g PL)+�����ӱ�����Լ���Triton X100,1g/l������50��C 30����ø������
���������IJ��Խ����ͼ15��16��ʾ��
|
|
|
ͼ15 ��ͬȥ�������账���Թ�����øЧ����Ӱ�� |
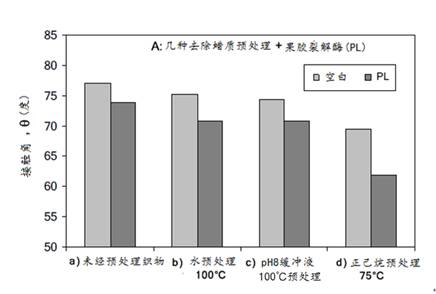
|
|
|
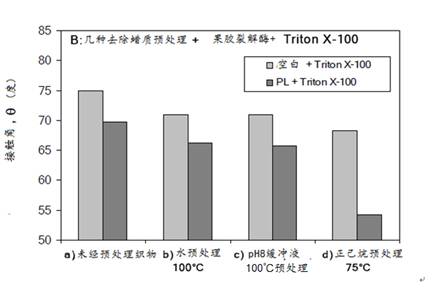
ͼ16��ͬȥ�������账����Triton X-100�ͽ��ѽ�ø������Ч��Ӱ�� |
1��A��������Ԥ�������հף��ͼ̺�������PL�������������ܽ���֯��Ľṹ�Ӵ��ǣ�����������Ԥ�����ͼ̺�PL�����Ľṹ�Ӵ����½������ԡ�
2��B����������Triton X-100��Ԥ�������հף��ͼ̺�������PL+ Triton X-100���������Ľṹ�Ӵ��Ǿ���A�鲻��Triton X-100���в�ͬ�̶��½�����������������Ԥ�����ͼ̺�PL�������½���������ֻ��55o����
Ϊ�˽�һ��֤ʵ���ʲ�ȥ��ȷʵ�����ڹ���ø�Թ�����ȥ�����ֽ����������ù���ø��������������Ԥ����+PL��������ķ�Ӧ���ʲ��ԣ������PL������Ϊ13 U/g ֯�����������ģ�����ȥ���ʣ���Ӧ����������ͼ17��ʾ��
|
|
|
ͼ17 �������账���Թ���ȥ���ʵ�Ӱ�� |
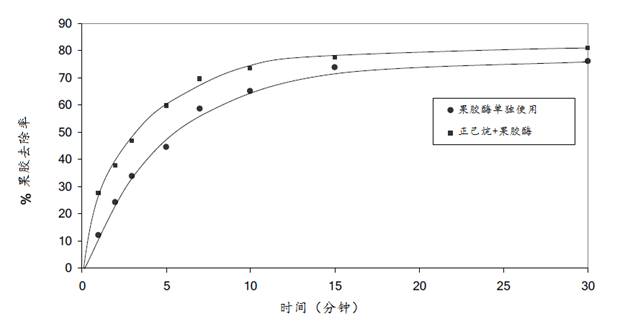
��ͼ17�ɼ�����Ӧ���ڵ����߳����ԣ�һ����Ӧ������һ����Ӧ���ߵ�63%������ȥ���ʣ���������ʱ�䳣����time constant*������������Ԥ����+PL������ʱ����5.5���ӣ������ù���ø����������Ҫ9���ӣ�ǰ�ߵķ�Ӧ�����Ǻ��ߵ�1.65������30���Ӻ�Ĺ���ȥ���ʷ��棬ǰ�߿ɴ�76-80%��������ֻ��70%������ʵ����֤�������ʲ��ȥ�������Լӿ����ø��ø�ٷ�Ӧ���ʡ���*63%�DZ���һ����Ӧ���ߵ�һ��������
����������ø��������Ҫ����
����øŨ�ȡ�pH���¶ȡ����ϼ��ͱ�����Լ�����ø��������Ҫ������
��1������øŨ������
ȡ10���˽���֯��ֱ���pH8��50mMTris-HCl����Һ����øŨ�ȴ�1U/g������20U/g��ø��Һ�У����º͵�ҡ����50oC����5���ӡ�Ȼ��ⶨÿ�鴦����֯��Ĺ���ȥ���ʣ���������øŨ�ȶԹ���ȥ���ʵ�������ߣ���ͼ18��ʾ��
|
|
|
ͼ18 ����øŨ�ȵ�Ӱ�� |
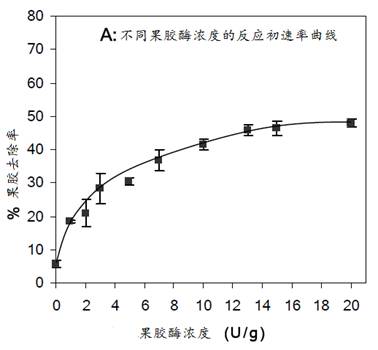
���������������øŨ����1U/gʱ�������ȥ����ֻ��6%,��Ũ��Ϊ5U/g��10U/g��13U/gʱ��ȥ���ʷֱ�����30%��41%��46%���������ӹ���øŨ�ȣ�������ˮ���������������ӡ�
��2�����pHѡ��
��������ͬ���������£�����ͬ������50mM�����Ρ�Tris-HCL�������λ���Һ����pH��6��7��7.5�� 8�� 8.5��9��9.5����10��PL����ø����Һ����֯�Ȼ��ⶨÿ�鴦����֯��Ĺ���ȥ���ʣ��Ƴ��������ߣ���ͼ19��ʾ��
|
|
|
ͼ19 PHֵ��Ӱ�� |
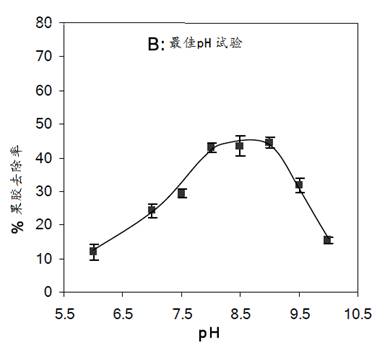
����ͼ�ɼ���������Һ��pH6���ö�������ˮȡ������Һ��ʱ������ȥ���ʽ�Ϊ11%����pH8-9ʱ����ȥ���ʿɴ�45%������PL�ĵȵ��Ϊ8.5��һ�¡�
��3�������¶�����
������¶ȷ�Χѡ����20oC��80oC��, ÿһ�������¶ȼ��10oC, Ȼ��ⶨÿ�鴦����֯��Ĺ���ȥ���ʣ������ͼ20��ʾ��
|
|
|
ͼ20 PHֵ��Ӱ�� |
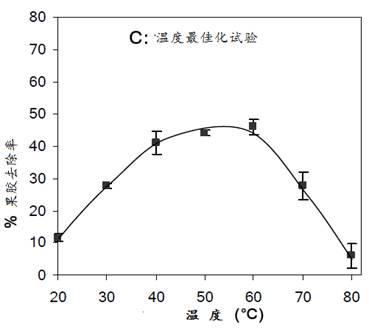
����ͼ�ɼ����¶ȶ�PLø�ٷ�Ӧ������������Ӱ�죬��20oC��30oCʱ������ȥ���ʽ�Ϊ15%��36%�����¶�����50oCʱ������ȥ���ʴﵽ��ֵ��46%����������60oCʱ���������û�б仯���˺����������70oCʱ�����ڸ���ʹPLøʧ�����ȥ�������½���27%��
��4�����ϼ�����Ӧ�ó����ѡ��
��ǰ�����������ᣨ�Σ��ѽ�ø��ø�ٷ�Ӧ������Ca2+������ԡ�м���EDTA���Ҷ��������ᣩ�Ƚ������ϼ�����ø�ٷ�Ӧ�к�ǿ���������á��������ѽ�ø��ø�ٷ�Ӧ������������֪������ά�ľ��Ʋ㣨������������ڼ䣩�У���Ҫ��δ�����������Ⱥܵ͵Ĺ�����������ˮ�����Թ������������и���ɣ���Ca2+�Ƚ����������к�ǿ�����������Ӷ�ʹ���ڵ����Թ������������γɽ�����ʹ���ԵĹ�������ӽ����һ����ɵ��νṹ���������ϼ����������νṹ�е�Ca2+�Ƚ��������ӣ��Ӷ�����ǵĽ�ϣ������ڹ�����ȥ���������������ϼ����ľ���Ч����������������������á����ھ���ԡ�к�������EDTA(0.5g/l)����ʹ�ѽ�øʧ���EDTA����ˮ����ʱ��ˮ�⣬����������һ���Ķ��ԡ��̺���������Ӱ����ٵ����������ƺ��ǰ��������������ࣨ��Bayer��˾����ƷBaysolex 21010�������ϼ������о����ߵ����ּ��뷽ʽ����1�����ϼ�����øͬԡ��2�����ϼ��ڹ���ø�������һ��95oC��ˮϴ��15���ӣ�ʱ���롣�����ͼ21��ʾ��
|
|
|
ͼ21 ���ϼ���Ӱ�� |
��ͼ�ɼ�����ͬԡ������ȣ��ڵ�һ����ˮϴʱ����2g/l
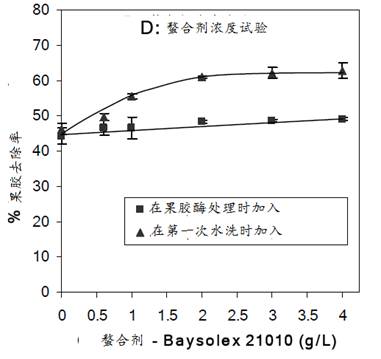
Baysolex 21010,��������߹���ȥ���ʣ���45%�����63%�����������Ϊ���ڼ��Թ����ѽ�ø�����������й����ȱ�ȥ���������ڵ�һ��ˮϴʱ�����ϼ���ȥ�����Ʋ��е��νṹ�е�Ca2+��ʹ����ĵ���������������������������ˮϴ�����Ӷ���һ������˹���ȥ���ʡ�
���ģ�����ø����ø�����Ͼ���
��ǰ��������������������������ȡЧ���������ڳ������й��������ø�Ӵ���������߹�����ȥ���ʡ���������������������Ԥ����������ʵ���Դˣ������������������о����ý���øԤ������/�������øͬԡ���������顣
����øԤ������Ĺ���ø����
����1��100U/g����ø����50mM Tris-HCl pH=8����Һ�У�30oC����15����
����2��13U/g�����ѽ�ø����50mM Tris-HCl pH=8����Һ�У�50oC����15����
5��������飺
A- ����1�Ͳ���2���Dz���ø�Ŀհ�
B- ����1����ø����+����2��������ø�Ŀհ�
C- ����1��������ø�հ�+����2����ø����
D- ����1����ø����+����2����ø����
E- ����1��1g/lTriton X-100�Ľ���ø����+����2����ø����
����������������������∼9710 N/m2�Ļ�е���á���ˮϴ���ﴦ�������ͼ22��ʾ��
|
|
|
ͼ22 �����Ͷ�����������Ч���Ƚ� |
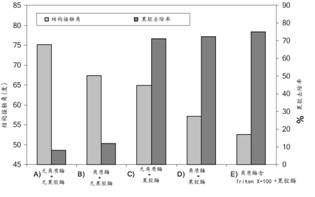
��ͼ�ɼ�����������ø��B��������ø(C)��������ʹ�ṹ�Ӵ��Ǵ�75o���͵���67o���ڹ���ȥ���ʷ��棬��B��������ֻ��9% ����(C)�����Ŀɴ�70%����D������øԤ����+����ø�ͣ�E���ڽ���øԤ����ʱ����Triton X-100������Լ������ģ�����ȥ����û�б仯�����ṹ�Ӵ�����ֱ���57o��51o��
���壩����ø����øͬԡ����
Ҫʵ�ֽ���ø����øͬԡ����ǰ�������ȳ������м������⣺
1�� ����ø����øͬԡ�����Ӱ��˴˵�ø�ٷ�Ӧ������ͨ������ø����øͬԡʱ������ø�Ļ������ԣ�֤�����������ݵġ�
2�� ��ǰ��֪������ø����ø�������¶ȷֱ�Ϊ30oC��50oC,������ø�ڣ�40oCʱʧ���������ͬԡ�������¶�����30oC,��Ҫ�������Ч����������߹���ø��Ũ�ȡ�
����ø����øͬԡ����������������50mM Tris-HCl pH=8����Һ�У�30oC����15���ӣ����������∼9710 N/m2�Ļ�е���á���ˮϴ���ﴦ����
���Ϊ��5�����飺
A�� �հ�
B�� ����ø����
C�� ����ø����
D�� ����ø����øͬԡ����
E�� ����1g/lTriton X-100��ͬԡ������
�����ͼ23��ʾ
|
|
|
ͼ23 ��ͬ����Ч��ˮƽ�Ƚ� |
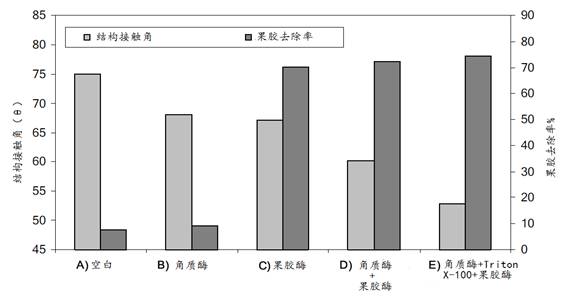
��ͼ23��֪������ø����øͬԡ����֯��Ľṹ�Ӵ����ȡ�60��,�����������ǵ��������ģ���ɽ���Ϊ����øȥ������ά��������ʣ������ڹ���ø�Գ����ڵĹ������ӿ��������������ά�������ʵ�ȥ�����Ӷ��������ˮ�ԣ�������1g/lTriton X-100��ͬԡ�����ĽӴ��ǿɴ���52����ˮ��ʪЧ����NaOH�����൱��
�塢����
1����֯Ʒ��ø�����¼������з�Լ�Ѿ�����15��࣬�������ⷽ��Ļ�����Ӧ���о���������ȡ���Ⱥ���ȡ�õĽ�չ��ֵ�����������ѧϰ�ͽ����Ŀǰ�ҹ�����ijЩ��֯Ʒ�ϵ�Ӧ����Щ���ܣ���Ȼ����ø���ƹ�Ӧ������Щ�����ģ�������������Ӧ���о�������δ��������ʵ�������������ƹ�Ӧ�õ��ֽš�����ֵ��ע������⡣Ŀǰ���ҹ���֯ҵ�������Լ800~900�֣�����Ʒ��Ⱦ���ӹ�������ľ���ø�Ƽ��Ĺ��������⣬ҲӦ�ǹ���ø�Ƽ���ҵ��ע������֮һ��
2��ø������Ⱦ������˵����Ⱦ�ϵ����ݣ�ʹӦ����Ա��������ĸо������ҳ��̵Ļ����ⶨ������������ͬ��ø��Ķ���Ҳ���ܸ��죬���״����Ʒ�����л��ȷ����Ϣ�������������Լ۱ȵ��жϡ�Ϊ�ˣ���˫���ܹ�ͬЯ������������Ȼ�Ľ��������
3����ø��Ӧ�÷��棬Ⱦ����ҵ�ĿƼ���ԱҪ�ӡ����ʹ��֮�����ɵ������ʹ֪֮������������������ɫ��֯Ʒø�������գ�Ը��ҵ��ͬ���ǹ���֮��
��Ҫ�ο����ϣ�
1��Agrawal P.B.-THE PERFORMANCE OF CUTINASE AND PECTINASE IN COTTON SCOURING, Thesis, University of Twente,the Netherlands,(2005)
2��Cristina M. L. Carvalho Cutinase structure, function and biocatalytic applications. Electronic Journal of Biotechnology Vol.1 No.3, Issue of December 15, 1998
3��Pasi Halonen��et al.,A HIGH THROUGHPUT PROFILING METHOD FOR CUTINOLYTIC ESTERASES��VTT BIOTECHNOLOGY
4) V. Brissos et.al Improving activity and stability of cutinase towards the anionic detergent AOT by complete saturation mutagenesis, Protein Engineering, Design & Selection vol. 21 no. 6 pp. 387�C393, 2008
5�������� ����ø�����ڷ�֯��ҵ�е�Ӧ�����й����﹤����־��2010��30��9����105��109
6��HACH LANGE Radiometer pH-Stat Theory and Practice
7) SongTao Xie (a Thesis for the degree of MS)Characterization of interyarn pore size and its distribution
8) TRI Auto-porosimeter
9) Danielle Biscaro Pedrolli et al. Pectin and Pectinases Production, Characterization and Industrial Application of Microbial Pectinolytic Enzymes. The Open Biotechnology Journal, 2009, 3, 9-18
10)��ѧ�ܵ� �����о��½�չ��Journal of Chinese Institute of Food Science and Technology�� Feb�� 2 0 1 0Vol�� 10 No�� 1
11)Pectinase Database-Specific activity pectinases enzyme activity enzymatic assay
12)Enzyme Explorer Assays by Enzyme Name Sigma-Aldrich
13��Dashek, William V., ed. Methods in plant biochemistry and molecular biology. Boca Raton, FL: CRC Press: pp. 309-321. Chapter 25.
14��Sigma Enzymatic Assay of PECTINASE (EC 3.2.1.15);Sigma Enzymatic Assay of PECTIN LYASE (EC 4.2.2.10)
15���л�������ҵ�� QB 1502-92 ʳ Ʒ �� �� �� �� �� ø �� ��
16)Atina.K.Losonczi (Ph.D.Thesis) Bioscouring of Cotton Fabrics
17)Emre Karapinar,et al. Scouring of Cotton with Cellulases, Pectinases and Proteases FIBRES & TEXTILES in Eastern Europe July / October 2004, Vol. 12, No. 3 (47)
18)��ǿ�� ��֯��ø����Ĺ����ʺ����ⶨ ���Ļ�ࣨ2003��11�£�
19)��ѩ�ٵ� ����ά��ʪ��Ӱ�����ط��� ��֯ѧ�� 2004 vol.25 No.1
20��Innova Biosciences:Enzyme units,Activity and specific activity explaned(��������)��
21��Miller et,al;Liquid porosimetry New methodology and applications, J.of Coloidal and Interface Science 163 163/170(1994)
22)Novozyme, Determination of pectin removal on Cotton by ruthenium red dyeing SOP NO:EUS-SM-0103, 02/01 1-6 (2003)