ø��ѧ����֪ʶ(��) yd12912
�Ϻ���֯��ѧ�о�Ժ
������(����)
ȫ��Ⱦ���¼���Ӧ���ƹ�Э���� ���鴦
����ø�ٷ�Ӧ�Ĺ���ԭ��
ø��һ�㻯ѧ����һ��,�����뻯ѧ��Ӧ,����Ӧǰ��û���ʺ����ı仯,Ҳ���ı���淴Ӧ��ƽ��㡣����һ�ɻ�ѧ������ͬ,ø�ǵ�����,�������������ӵĸ������ԡ�
ø�ٷ�Ӧ�ġ��м���ѧ˵:
��ø�ٷ�Ӧ��,���dzƷ�Ӧǰ�ķ�Ӧ��Ϊ�����Ӧ��Ϊ���
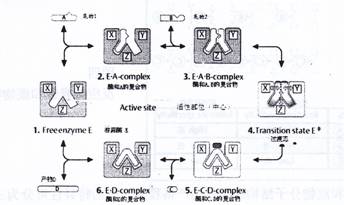
ø(E)���������(S)�����м�������ø-���︴���ES)����Ȼ���⸴�����ٷֽ�ɲ��P���������ø��E�����������ͼ����ʾ
|
|
|
ͼ�� ˫����ø�ٷ�Ӧʾ��ͼ |
(һ)ø�ٷ�Ӧ������
(1)ø�м��ߵĴ�Ч��
һ�������,ø�Ĵ�Ч�ʱȷǴ���Ӧ��108-20��,�ȷ�ø����Ӧ��107-13����
�ڻ�ѧ��Ӧ��,ֻ�е���ײ�ķ����������ڻ�ijһ��������Ec(���)�Ļ����ʱ,������Ч��ײ,�ŷ�����ѧ��Ӧ��ʹ���Ӵӳ�̬ת��Ϊ��������ѧ��Ӧ�Ļ�Ծ״̬����Ҫ��������Ϊ��ܡ���ͼ����ʾ
|
|
|
ͼ�� ø�ٷ�Ӧ�����л�ܵĸı� |
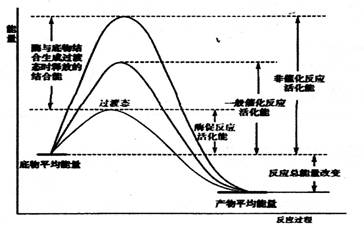
ø�ٷ�Ӧ����Ҳ�����ܵĴ�С�������,���Խ��,��Ӧ����Խ��,��˽��ͻ�ܻ���Ч�شٽ���Ӧ�Ľ��С�
(��)ø�ٷ�Ӧ�������
����ø�ٷ�Ӧ���ں��º͵�������˳�����ٽ��С�ͬʱ�Է�Ӧ����(���¶ȡ�pH��)���ϸ��ѡ������,���Ե���ͷ�Ӧ����Ҳ���ϸ��ѡ���ԡ���ͼ����ʾ:
|
|
|
ͼ�� ��Ӧ�����Ժ͵��������� |
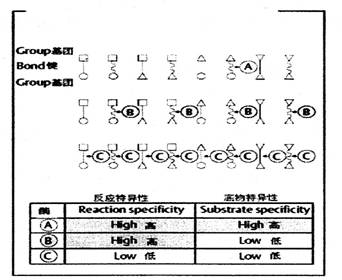
����ø�Ի�ѧ��Ӧ���ͺ͵�����ӽṹѡ���Ե��ϸ�̶�,ø�������Կɷ�Ϊ��������:
(1)���������ԣ�absolute specifity������ͼ����øA�Ե���ͷ�Ӧ���иߵ�������,Ҳ����˵,øAֻ��������һ�ֵ���,�����ض��Ļ�ѧ��Ӧ,�����ض��ṹ�IJ�����������Գ�Ϊ���������ԡ�
(2)���������(relative
specificity)������һЩø����������Խϲ�,��Щøֻ��������һ������һ�ֻ�ѧ��,��ͼ���е�Bø�����ֲ�̫�ϸ��ѡ����,��Ϊ��������ԡ�
(3)�����칹�����ԣ�sterospecificity����������ø�ܱ����ﲻͬ�������칹��,ֻ������һ�ֹ���������á����������칹�����Ժͼ����칹�����ԡ�
����)ø�Ļ������ģ�active
center����ƻ��Բ���active site��
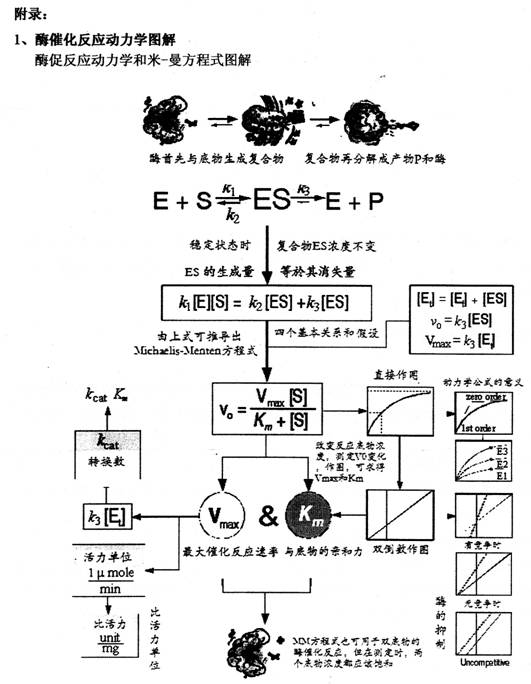
�������ø�ǵ�����,���ɸ��ְ�����������ļ����ӵ���״����ӡ����ڷ�������-NH2��-COOH���ź��ֲ���,���Ǽ��������������Ӽ��ͷ��»�������,����ø�й��۽��,������,��ʹ�����ʷ��ӳʡ����š�״(�����ṹ-tertiary structure)����ͼ����ʾ
|
|
|
ͼ�� |
ø�Ļ���������ø�͵������ɸ����������������Ӧ�IJ�λ��
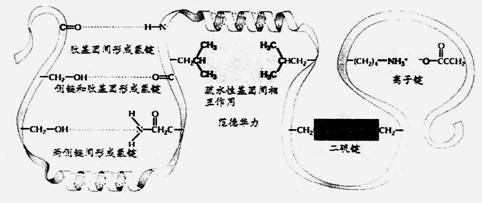
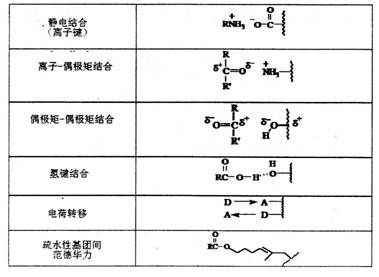
�ڻ���������,ø�͵������ͨ�������ż�ķǹ��۽��,����ø-���︴������1��ʾ:
|
��1 ø�͵����ķǹ��ۼ����� |
|
|
��һ������ø��˵,����������ָ�ڡ����š�״��ø���ṹ��������������һЩ������л�����л��ϵ�ijЩ������ɵ����������������������ţ��D�����������(��ø)����γ�ø-���︴����Ľ�ϻ���(binding group)����һ����Ӱ�������ijЩ��ѧ���ȶ��ԵĴ�����(Catalytic
group),����ʹ�����ľɼ��������ѡ��γ��¼�,��������ת��Ϊ�������һЩ��Ҫ��ø��ø��˵,��ø���������������ijЩ���š��ṹ���ǻ������ĵ���ɲ��֡�
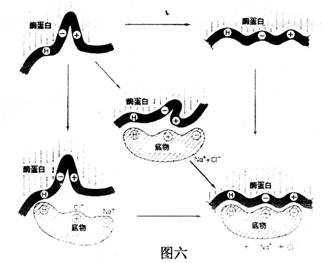
����,ø�����л���һЩ��������,������Ȼû�н�Ϻʹ�����,����Ӫ��һ��ʹø�Ļ������Ĵ��ں��ʵĿռ乹��,�������Ѩ���ѷ�,�յ������������Žӽ��ͽ��,���˳���Ϊ�յ�����(induced fit),��ͼ����ʾ��
|
|
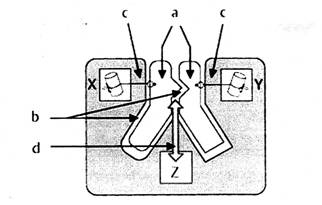
����Ӧʱ,ø�Ļ��������Ⱥ���У�a.�����ø����(approximation)����λȡ��(orientation)�յ����ϣ�b.�ų������ܼ�����,��ˮ,ʹ������ø�������Ӵ���c.����̬���ȶ�����d.����ת�����ĸ����̡���ͼ����ʾ��
|
|
|
ͼ�� |
(��)ø�Ļ���(activity)
ø�Ļ�����λ:
(1)���ʵ�λ(IU):
1961�����øѧ��(the international Union of Biochemistry)���,I964�걻���á����壺1��ø������λ��ָ���ض�����(
*ע��:�ڹ��ʶ�����(CGPM)��,SI)��ʱ�䵥λ����(s)���Ƿ���(min)��
(2)Kata1��λ(Kat):
��������ì��,1978����ʶ�����(CGPM)�Ƽ���1999��ٷ�����,��KatalΪø��������λ,ָ���ض������¡�ø�����ﱥ��ʱ,ÿ����ת��1mo1����������ø��,
1IU=1/6O ��katal=16.67n kata1
ø�ıȻ���(specific
activity)��
ÿ����ÿ����ø������
(3)ת����(Turnover
nunber)Kcat
��øѧ��,���嵥λʱ�䡢ÿ��ø�����л��Բ�λ��ʹ����ת���ɲ������������,���ɰ��������-������ʽ, Kcat=Vmax/[E]T�������,ʽ��[E]TΪ��(total)��øŨ�ȡ�
����ø�ٷ�Ӧ����ѧ
ø�ٷ�Ӧ����ѧ���о�ø�ٷ�Ӧ�����Լ��������ضԷ�Ӧ���ʵ�Ӱ�졣��ЩӰ�������У�����Ũ�ȡ�ø��Ũ�ȡ�pH���¶ȡ����Ƽ��Լ�������ȡ�
��ø�ٷ�Ӧ��,ø��E����������S)���ɡ��м�����ø-���︴����(ES)��,Ȼ���⸴�����ٷֽ�ɲ��P���������ø��
![]()
(һ)����Ũ��[S]��ø�ٷ�Ӧ��ʼ����(V)��Ӱ��
��1����-������ʽ(Michaelis-Menten Equation)����-������Km(Michaelis-Menten
Constant)
l913��Leonor
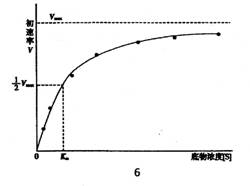
Michaelis��Maud L.Menten�������м���,�Ƶ�����-������ʽ,����ʾ����Ũ��[s]�仯��ø�ٷ�Ӧ��ʼ����(V)Ӱ��ķ���ʽ(����ʽ�Ƶ�����)��
��Щ�����ǣ�(1)�����ﷴӦ��(2)�ⶨ�ķ�Ӧ�����dz�ʼ����(initial rate),��Ϊ��Ӧ�ոտ�ʼ,���ɲ���������,������淴Ӧ�ɺ��Բ��ƣ�(3)�ڷ�Ӧ��ʼʱ,����Ũ��[S]ԶԶ����øŨ��[E],[S]�仯���Ժ��Բ��ơ�
![]()
V-��ͬ����Ũ��[S]ʱ�ij�ʼ����
Vmax ��������ʼ��Ӧ����
Km ������-������,�dz�ʼ����VΪ1/2 Vmaxʱ�ĵ���Ũ��[S]��
Kl��K2��k3�ֱ�ΪøE�͵���S���ɸ�����ES����������淴Ӧ������ֽ����ɲ��������ø���������ʳ���
����-������ʽ�����۳���
l��������Ũ��[S�ܵ�ʱ,��[S]����Km,ʽ�з�ĸKm+[S]��Km,��V=Vmax/Km,��Ӧ����V��[S]������(һ����Ӧ,���������);
2�������Ũ��[S]�ܸ�,��[S]����Km,Km+[S]��[S],��V��Vmax,��ʱ����ø�����еĻ��Բ�λ�������ﱥ�͡�
3����V=1/2
Vmax,���뷽��ʽ,��Km=[S],����ʾ����Ӧ����Ϊ������ʵ�һ��ʱ,��-������Kmֵ���ڵ���Ũ��[S]��
(2)����������Km)
��-������(Km)��ø��������������һ����Ӧ����(�¶ȡ�pH������ǿ��)��,��ֻ��ø������͵���������й�,��ø��Ũ���ء�
��ø�������ֲ�ͬ�ĵ���,��ÿ�ֵ��ﶼ��һ���ض���Kmֵ,����Kmֵ��С�ijƸ�ø�����ʵ��KmֵС,Ҫ�ﵽ��Ӧ����Ϊ������ʵ�һ��ʱ,����ĵ���Ũ��[S]�͵͡�
��������ES����E��S������K2ԶԶ����ES�ֽ������øE�Ͳ���P������k3ʱ,��K2����K3,K2+��3�Ԧ�2,��
|
Km= |
|
= |
������ES����E��S�����ʳ��� |
=Ks����ⳣ���� |
|
K1 |
ø�͵������ɸ�����ES�����ʳ��� |
��ʱKm����ø��������������ʵ����,�������е�ø�ٷ�Ӧ��,��Զ��k2����k3,����Km��Ks(��ⳣ��)����ͬһ������,�����ȡ��ʹ�á�
KmֵԽ��,ø���������ԽС,Ҫ�ﵽ������ʵ�һ��ʱ�������Ũ��[S]Խ��:��֮, KmֵԽС,ø���������Խ��,��Ҫ�ﵽ������ʵ�һ��ʱ�������Ũ��[S]�͡���ͬ�����ø,���ض��ĵ���,�в�ͬ��Kmֵ������һ��δ֪��ø,���ڹ涨�������²ⶨ��Kmֵ���Լ�����
(3)����ͼ���ⶨ���ø��Vmax��Kmֵ,��ͼ����ʾ
1����-����ʽ����
|
|
|
ͼ�� |
��ͼ����ʾ,��-������ʽ����������������ɣ�
��һ���֣���ʼ��Ӧʱ��ֱ�߲��֡���ʱ��Ӧ���ʺ͵���Ũ��[S]������,��Ӧ��һ����Ӧ��
�ڶ����֣���ߵ���Ũ��,ʹø���ӵĻ��Բ�λ����ռ��,ͼ���в�������,��ʱ��Ӧ����ϼ���Ӧ��
�������֣�������Ũ�Ⱥܸ�,ʹ���е�ø���ӵĻ��Բ�λ��������ռ�кͱ��͡���ʱ,�����ӵ���Ũ��,��Ӧ���ʱ��ֲ���,�ɽ�����,��Ӧ���㼶��Ӧ��
2��Lineweaveer-Burg(�����-��)˫������(double reciprocal plot)��ͼ��Kmֵ��Vmax��
������,�����ö��ַ����ⶨ��-������Km��Vmaxֵ������,�ڷ�Ӧ�й̶�ø��Ũ��,�ⶨ�ڲ�ͬ����Ũ���µij�ʼ��Ӧ����,���Km��Vmaxֵ������������-������ʽ�����ڽӽ�Vmaxʱ�ǽ�������,���Ѳ����ʵ��Vmax,���Բ���ֱ������ʼ���ʶԵ���Ũ����ͼ�����Km��Vmaxֵ��
1930��Lineweaver��Burk����-������ʽ����ȡ����,������ʽ����ʽ��ʾ1/V��1/[S]��ֱ�߹�ϵ,ʽ��,Km/Vmax��б��,1/Vmax��y���ϵĽؾࡣ
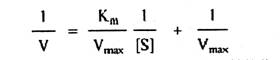
��1/V��l/[S]��ͼ,���ɵõ�һ��ֱ�ߡ���ֱ����Y��Ľؾ�Ϊ1/Vmax,��X���ϵĽؾ�Ϊ-1/Km�ľ���ֵ(��ͼ����ʾ)��
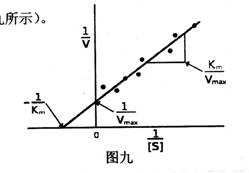
(��)�ڵ���Ũ���㹻��ʱ,��Ӧ������øŨ����߶���ֱ������
����Ӧ��ϵ��,����Ũ�Ⱥܸ�,�Ҵ��øŨ��,ø���������л��Բ�λ����������ɸ�����ES,�ұ����ﱥ��ʱ,��Ӧ���ʣ�V=K3[ES]=K3[E]���ﵽ���ֵVmax����ʱ,��Ӧ��һ����Ӧ,����Ӧ������øŨ�����ӳ����ȵ�ֱ�߹�ϵ,��ͼʮ��ʾ��
|
|
|
ͼʮ øŨ�ȶ�������ͼ |
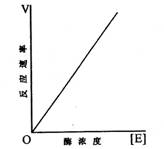
��)�¶ȶ�ø�ٷ�Ӧ����Ӱ���˫����
ø���������,�¶ȶ�ø�ٷ�Ӧ��˫��Ӱ�졣�����¶�һ������Լ���ø�ٷ�Ӧ���ʡ�����,��Ϊ�����ø�ǵ����ʴ����,��̬��,��������и��ֻ��ŵ������,ʹø����������ȶ��ġ����š�״,���������ľ��������ŵ���Ѩ���档����¶Ȼ��ƻ����ż�������,����ʱ��ʹø����ʧ��,���Թ�������¶ȷ���ʹø����ʧ��,��������ת��
�����ø���¶�
ø�������¶Ȳ���ø�����Գ���,��Ϊ���淴Ӧ���е�ʱ���йء�ø�����ڽ϶�ʱ���ڳ��ܽϸߵ��¶ȡ��෴,���ŷ�Ӧʱ���ӳ�,���ʷ�Ӧ�¶�Ҳ�ή�͡�������ή��ø�Ļ���,�������ƻ�ø,���¶Ȼ�����,ø�Ļ����ֻ�ָ���
(��)pH��ø�ٷ�Ӧ���ʵ�Ӱ��
ø���������Եĸ߷��ӵ����,�ڲ�ͬpH������,ø���ӵ����Ի��źͼ��Ի��Ž�������ͬ��������ر��Ǵ��ڻ������ĵĻ���),ֻ�е���Щ���Ŵ���һ�������״̬��,�ŶԵ���������������ͬʱpHҲ��ı�ijЩ����(�����ʡ��������)��ø�����̶�,�Ӷ�Ӱ��������ø�͵���Ľ�ϡ�
�����ø�Ļ��������ܻ���pH��Ӱ�졣ֻ����ijһpHʱ,ø�ķ�Ӧ�������,����ø������pH(optimum
pH)�����ڻ���ڴ�pH,��Ӧ�����½�����ͼʮ����ʾ
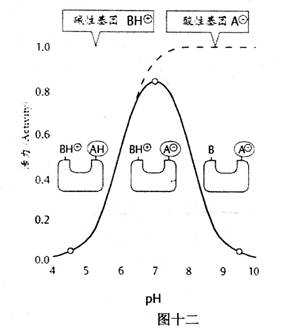
����pH����������ࡢŨ�Ⱥͻ�����ϵ��ͬ����ͬ,���ҳ���ø�ĵȵ�㲻һ�¡��������pH����ø����������,ֻ����һ�������²������塣pH���ͻ���߶���Ӱ��ø�Ĺ���,��������ø�ı��Ժ�ʧ�
(��)���Ƽ��ͼ�����Է�Ӧ�ٶȵ�Ӱ��
��1�����Ƽ�
һЩ����ø�ı����������,�Ӷ�����ø�Ĵ����Ե�������ø���ױ��Ե�����ͳ��Ϊø�����Ƽ�(inhibitor)���������Ƽ���ø�������ĵı�����Ž�ϵĽ��̶ܳ�,���Ƽ���Ϊ������������(irreversible inhibition)�Ϳ�������������(reversible
inhibition)
1)��������������(irreversible inhhibiton)
���Ƽ���ø�������ĵı�����Ź��۽��, ʹøʧ��,�Ҵ����Ƽ��������������˵ȷ�������ȥ����
2)����������������(reversible inhibition)
�������Ƽ�ͨ���Էǹ�����ø��(��)ø-���︴��������Խ��,ʹø���Խ��ͻ���ʧ�����������˵ķ����ɽ����Ƽ���ȥ��������������������ֿɷ�Ϊ��������:�������������á��Ǿ������������úͷ��������������á�
(2)�������(activator)
��ʹ����øת��Ϊ�л��Ի�ʹø�������ӵ����ʳ�Ϊø�ļ��������������Ϊ��������,��Mg2+,K+,Mn2+��,����Ϊ������,��Cl-�ȡ�Ҳ�������л������D���,�絨֭���εȡ��������ø��������һ��ѡ����,�����ø�ٷ�Ӧ����Ҫ�̶�,������ֿɷ�Ϊ���輤���(essential activator)�ͷDZ��輤��� (non-essential activator)
2�������ʽṹͼ
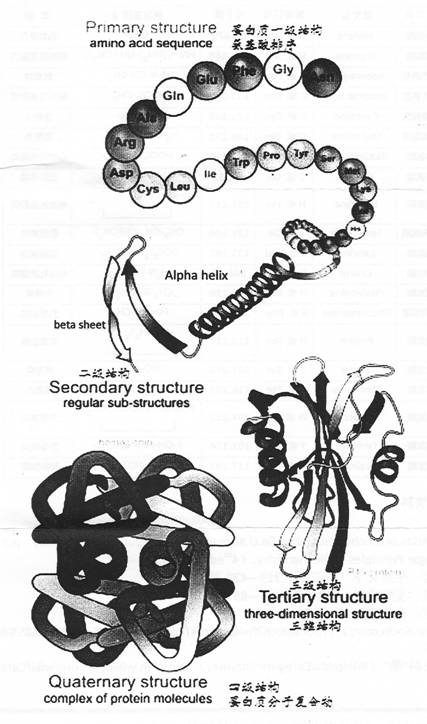
3)20�ְ�����
��Ҫ�ο�����
[1]Color
Atlas of Biochemistry ,.2nd.Ed.(J.koolman,K.H.Roehm)(Thleme,2005)p88-110
[2]Lehninger Principles of Biochemistry.(4-edition��p190-237
[3]������ ���ﻯѧ(������)��31-429
[4]������ ���ﻯѧ(���߰�)��64-85
[5]������:�йء�ø��ѧ������http://www.docin.com/app/docsearch?fn=&searchType_banner
=p&dky=%E9%85%86%E5%
[6]ά���ٿơ�ø��(WikipediaCategory:Enzymes) http://en.wikipedia.org/wiki/Category:Enzymes